A Synopsis of ‘The Seven Sins of Evolutionary Psychology’
Contents
Evolutionary Psychology: An Exchange
| Jaak Panksepp/Jules B. Panksepp | 2 | A Synopsis of “The Seven Sins of Evolutionary Psychology” |
|||
| Ullica Segerstråle | 6 | Minding the Brain: The Continuing Conflict about Models and Reality |
|||
| Linda Mealey | 14 | “La plus ça change…”: Response to a Critique of Evolutionary Psychology |
|||
| Carlos Stegmann | 20 | Comments on Jaak Panksepp and Jules B. Panksepp’s Paper “The Seven Sins of Evolutionary Psychology” |
|||
| Russell Gardner, Jr. | 25 | Affective Neuroscience, Psychiatry, and Sociophysiology | |||
| Gerhard Meisenberg | 31 | Degrees of Modularity | |||
| Ian Pitchford | 39 | No Evolution. No Cognition | |||
| Scott Atran | 46 | The Case for Modularity: Sin or Salvation? | |||
| Jaak Panksepp/Jules B. Panksepp | 56 | A Continuing Critique of Evolutionary Psychology: Seven Sins for Seven Sinners, Plus or Minus Two |
General Articles
| Shulamith Kreitler | 81 | An Evolutionary Perspective on Cognitive Orientation | |||
| Chris MacDonald | 98 | Evolutionary Ethics: Value, Psychology, Strategy and Conventions |
|||
| 106 | Zusammenfassungen der Artikel in deutscher Sprache |
Impressum
Evolution and Cognition: ISSN: 0938-2623 Published by: Konrad Lorenz Institut für Evolutions- und Kognitionsforschung, Adolf-Lorenz-Gasse 2, A-3422 Altenberg/Donau. Tel.: 0043-2242-32390; Fax: 0043-2242-323904; e-mail: sec@kla.univie.ac.at; World Wide Web: http://www.univie.ac.at/evolution/kli/ Editors: Rupert Riedl, Manfred Wimmer Layout: Alexander Riegler Aim and Scope: “Evolution and Cognition” is an
interdisciplinary forum devoted to all aspects of research on cognition in animals and humans. The major emphasis of the journal is on evolutionary approaches to cognition, reflecting the fact that the cognitive capacities of organisms result from biological evolution. Empirical and theoretical work from both fields, evolutionary and cognitive science, is accepted, but particular attention is paid to interdisciplinary perspectives on the mutual relationship between evolutionary and cognitive processes. Submissions dealing with the significance of cognitive research for the theories of biological and sociocultural evolution are also welcome. “Evolution and Cognition” publishes both original papers and review articles. Period of Publication: Semi-annual Price: Annuals subscription rate (2 issues): ATS 500; DEM 70, US$ 50; SFr 60; GBP 25. Annual subscriptions are assumed to be continued automatically unless subscription orders are cancelled by written information. Single issue price: ATS 300; DEM 43; US$ 30; SFr 36; GBP 15 Publishing House: WUV-Universitätsverlag/Vienna University Press, Berggasse 5, A-1090 Wien, Tel.: 0043/1/3105356-0, Fax: 0043/1/3197050 Bank: Erste österreichische Spar-Casse, Acct.No. 073-08191 (Bank Code 20111) Advertising: Vienna University Press, Berggasse 5, A-1090 Wien. Supported by Cultural Office of the City of Vienna, Austrian Federal Ministry of Science/ Transportation and the Section Culture and Science of the Lower Austrian State Government.
A Synopsis of “The Sevens Sins of Evolutionary Psychology”
Our target article (PANKSEPP/PANKSEPP 2000) summarized some common and uncommon concerns that neuroscientifically oriented investigators of the mind/brain may have with currently fashionable versions of evolutionary psychology. Our main thesis was that an evolutionary psychology that seeks to understand “human nature” will be woefully incomplete and perhaps misguided if it does not incorporate the lessons from the past century of research on subcortical emotional and motivational systems that all mammals share. It is advisable to begin building an evolutionary viewpoint of mind based on comparative concepts that incorporate the intrinsic, evolutionarily provide systems found in all mammalian brains.
The more evangelical varieties of psycho–evolutionary thought that arose from social-science traditions have typically not been concerned with how the human mind emerges from specific brain functions. Thus, our argument was motivated by three converging and overlapping issues: (1) The failure of evolutionary psychology (e.g., BUSS 1999, and many others) to address the evolved, adaptive mechanisms in the mammalian brain that have been and are being studied by many investigators in the neurosciences. (2) The prolific tendency of evolutionary psychology to generate adaptive stories, especially with regard to human socio–emotional processes, without much apparent concern about how such presumed adaptations might be instantiated in neural systems through a great diversity of brain–environment interactions (e.g., COSMIDES/TOOBY 2000). (3) The concurrent claim that evolutionary psychology is now providing one of the most robust lines of thought for identifying the types of evolved mechanisms that actually exist in the human brain (TOOBY/COSMIDES 2000).
Our discussion was framed with the recognition that several distinct schools of thought are currently providing conceptual frameworks to help guide future empirical inquiries in the ongoing attempt to understand how evolution constructed the mind/ brain of humans and other animals. Also, our article was premised on the full acceptance that evolutionary viewpoints are essential for specifying the neuropsychological natural kinds that actually exist, as ancestral birthrights, within the brains of humans as well as other animals.
Our specific concerns were as follows:
1. Are there really Pleistocene sources of current human social adaptations? Evolutionary Psychologists seem to be backing off from this assumption as they begin to realize that neocortex is the main type of cerebral tissue that emerged two million years ago, when hominid brains diverged from the “chimp-sized” brains of our ancestral stock. Thus, the critical question is whether special-purpose programs emerged in neocortical tissues primarily through genetic changes that occurred in the human line during the past few million years. That issue probably cannot be answered unambiguously without concurrent behavioral neuroscience and molecular biology research. If such adaptations emerged, they are likely to reflect quite general information handling capacities, permitting abilities ranging from art to human languages, from thought to culture. Most evolved, special-purpose systems may be devoted to functions we still share with other animals.
2. Excessive species-centrism in Evolutionary Psychology. Completion of the human genome project has unequivocally highlighted the likelihood for genetic similarities across all mammalian species. We suggest that without a clear recognition of what we share with other animals, it will be impossible to specify what is truly unique in the intrinsic, genetically-provided capacities of the human mind/brain. The neuroanatomical and neurochemical homologies that exist in all mammalian brains presently provide the most substantive way to specify the types of evolved faculties in our own minds.
Evolution and Cognition ❘ 2 ❘ 2001, Vol. 7, No. 1
3. The sin of adaptationism. It is well recognized that one must always discriminate between adaptations arising from natural selection and those arising from the emergence of flexible abilities to learn new skills and strategies. The cortex obviously becomes specialized for a large number of special purpose skills and abilities during development (many of which may seem superfically “encapsulated” because most of our thoughts are constrained by primitive emotional systems that regulate our feelings), but current evolutionary psychology provides no special inroads to understanding which adaptations have truly emerged through natural selection.
4. The sin of massive modularity. From a neuroscience perspective, “modularity” is an obsolete concept, resembling the “centers” concept that was discarded by scientists doing brain research several decades ago. There are many special-purpose circuits and systems in subcortical areas of the brain, but those systems are not encapsulated and appear to interact with each other extensively. For instance, during every emotion, the activities of widespread brain areas are modified. Obviously, a critical scientific issue is the degree to which the specialized functions of higher cerebral abilities exist as birthrights and how much they reflect the operation of developmental programs.
5. On the conflation of emotions and cognitions.
The cortex mediates many specialized cognitive abilities. Most of the key systems for mammalian emotionality are subcortically organized, and the neglect of these systems can only lead to a very mistaken view of how emotionality is truly elaborated in the human brain. Disciplines that focus simply on the higher cortico–cognitive aspects of emotional processing are bound to make many mistakes about the fundamental nature of emotions and affective processes (PANKSEPP 2001).
6. The absence of credible neural perspective in Evolutionary Psychology. This is a general criticism of all mind sciences that fail to take real brain issues into consideration. Although the full potential of the mind emerges from organismic–environmental interactions, all foundational issues need to be linked to our gradually emerging understanding of how specific neural systems function. The neuroscience revolution has provided enough raw evidence for everyone to begin thinking more clearly about those underlying brain issues.
7. Anti-organic bias or the computationalist/representationalist myth. There have always been two grand perspectives for explaining the organization of mind—mind as an organic process and mind as a “spiritual” process that can exist independently of the material world. The modern computational view still tends to subscribe to a modern variant of the latter view. From a non-dualistic, materialistic perspective, that may be highly misleading. All minds are probably strongly constrained by the organic–neural processes by which they are instantiated. To neglect those issues, in preference for a strict computationalist view, is likely, we believe, to retard deep understanding. In our reply to the commentators, we highlight a recent critique by FODOR (2000), one of the great promoters of the computationalism, who now agrees that grave errors have been made during the cognitive revolution.
Our criticisms do not apply equally to all existing lines of psycho–evolutionary thought, but mainly to the rhetorically most audible social psychological variants that arose from the ashes of the contentious “sociobiology wars” of the late 1970s and early 80s. Additional issues could have been raised, such as the massive interactions of certain well-characterized brain systems (e.g., dopamine, norepinephrine, serotonin, acetylcholine, GABA and glutamate) which participate heavily in all of the processes about which evolutionary psychologists opine. Several related “sinful” tendencies could have been highlighted, but they all ultimately relate to the failure of Evolutionary Psychology to assume a deeply organic stance that fully respects the role of developmental systems in the epigenetic creation of higher mental faculties.
Our overall view is that the developmental interactions between ancient, special-purpose circuits and more recent, general-purpose brain systems, such as those found in the neocortex, can generate many of the “modularized” human abilities that Evolutionary Psychology has entertained. By simply accepting the remarkable degree of neocortical plasticity, especially during human development, genetically-dictated, sociobiological “modules” begin to resemble products of dubious human ambition rather than sound scientific inquiry. We believe that if Evolutionary Psychology attempts to construct a view of human nature based upon inclusive-fitness issues and improbable neurological assumptions, while continuing to avoid a substantive confrontation with our ancient animalian heritage, it is as likely to retard lasting scientific progress as to advance it. In short, the only place many concepts of
Evolution and Cognition ❘ 3 ❘ 2001, Vol. 7, No. 1
evolutionary psychology may be encapsulated are in the minds of scholars who, all too often, have little wish to immerse themselves in the essential neuroscientific and genetic issues.
We recognize that there are recent strands of thought in evolutionary psychology that take positions more congenial to our own. For instance, we admire BADCOCK’s (2000) confrontation with key biological issues, as well as his willingness to try to link evolutionary thought to earlier intellectual traditions. We especially appreciate the way he handles the possibility that cultural issues, obviously important in their own right, may be linked to some important heritability findings such as genomic imprinting. Stunning discoveries such as that of KEV-ERNE and colleagues (1996), indicating that the development of neocortex may be influenced more by maternally-imprinted genes while the development of key emotional areas such as the hypothalamus are governed more by paternally-imprinted genes, provide novel ancestral–genetic tethers over brain organization that may have profound implications for the nature–nurture controversy.
Of course, we remain as intellectually stimulated by evolutionary psychological stories as anyone. We have been especially entertained by the recent willingness of some theorists to recognize the importance of the emergence of artistic endeavors in the socio–cortical emergence of our species (MILLER 2000) and of course, the view of language as partly a social-grooming adaptation (DUNBAR 1995) which fits nicely with some animal social-systems data (PANKSEPP 1998a). However, we would again advise evolutionists to view the neocortex as the generalpurpose playground for our basic attentional, emotional and motivational systems, as opposed to their sources. Indeed, it may well be that such a generalpurpose associational spaces are the ideal playgrounds for the many dimensions of human creativity and entertainment, both abundantly serious and humorous depending on the depth of our emo-
tional personalities as much as anything else. With our neocortex we can do whatever we can imagine—and because of the mass of general purpose associational space that we possess, there are few obvious limits to our imagination. This affords us great liberties in our humanistic endeavors and helps create great havoc for our scientific ones.
We would simply note that the expansion of instincts at the turn of the previous century is now being matched by the postulation of genetically modularized cognitive functions. The “expansion of instincts” failed scientifically because they were not tethered to brain systems, and the expansion of “modularized” functions may have a comparable fate for similar reasons. A recent, and more guarded attempt to bring instinctual systems back to the forefront of our thinking (e.g., PANKSEPP 1998a) has been premised on only entertaining basic psycho– behavioral entities for which there exist robust and converging neuroscientific evidence. This also gives us considerable confidence in defining what types of systems we need to study more earnestly in humans, while preventing mere human creativity from being the major guide of what does or does not exist in the natural organization of our minds. If we carefully work out the fundamental neuro-psychological processes that we share with other animals, we will be in a better position to comprehend what is truly unique in our own mind/brains.
Our implicit aim in the target article was to strongly encourage similar constraints in evolutionary psychological thought. Obviously, there are many stories that remain to be told around this new intellectual campfire, and the likelihood that primitive forms of affective self-representation still link us to a deep animalian past may be a worthy concept for mainstream thought in evolutionary psychology (PANKSEPP 1998a,b). Indeed, this view may allow us to better understand and harmonize scientific and humanistic endeavors. We do hope that an increasing number of scholars will begin to incorporate the evolutionary passages of deep-time into their considerations of the evolved nature of human mind-flesh… most especially in ways that can lead to the deep empirical evaluation of ideas. Being inheritors of a vast and to some extent, general-purpose cerebral canopy,
References
- Badcock, C. (2000) Evolutionary Psychology: A Critical Introduction. Polity Press: Cambridge UK.
- Buss, D. M. (1999) Evolutionary Psychology: The New Science of the Mind. Allyn and Bacon: Boston.
- Cosmides, L./Tooby, J. (2000) Evolutionary psychology and the emotions. In: Lewis, M./Haviland, J. (eds) The Handbook of Emotions, 2nd edition. Guilford: New York, pp. 91–116.
- Dunbar, R. (1995) Grooming, Gossip and The Evolution of Language. Harvard University Press: Cambridge, MA.
- Fodor, J. (2000) The Mind Doesn’t Work That Way, The Scope and Limits of Computational Psychology. The MIT Press: Cambridge, MA.
- Keverne, E. B./Fundele, R., et al. (1996) Genomic imprinting and the differential roles of parental genomes in brain development. Developmental Brain Research 92: 91–100.
Miller, G. (2000) The Mating Mind: How Sexual Choice
Shaped the Evolution of Human Nature. William Heinemann: London UK.
- Panksepp, J. (1998a) Affective Neuroscience, The Foundations of Human and Animal Emotions. Oxford University Press: New York.
- Panksepp, J. (1998b) The periconscious substrates of consciousness: Affective states and the evolutionary origins of the SELF. Journal of Consciousness Studies 5: 566–582.
- Panksepp, J. (2001) At the interface between the affective, behavioral and cognitive neurosciences. Decoding the emotional feelings of the brain. Brain and Cognition, in press.
- Panksepp, J./Panksepp, J. B. (2000) The seven sins of evolutionary psychology. Evolution and Cognition 6(2): 108– 131.
- Tooby, J./Cosmides, L. (2000) Toward mapping the evolved functional organization of mind and brain. In: Gazzaniga, M.S. (ed) The New Cognitive Neuroscience, 2nd edition. MIT Press: Cambridge MA, pp. 1167–1178.
Minding the Brain
The Continuing Conflict about Models and Reality
HERE IS THE FAMOUS joke about a traveler asking a local if he knew how far it was to a particular town. Yes, said the man. But, he added, you can’t get to there from here! This may be rather like the way Jaak and Jules PANKSEPP view the prospect of elucidating the mind with the help of evolutionarily informed cognitive neuroscience. Theirs is one of the strongest scientific objections so far to the whole enterprise of evolutionary psychology. For the PANKSEPPs, there is only one correct way to go, and that is through the field of affective neuroscience. You cannot find out about the functioning of the mind/brain by asking questions about adaptation and design, and by reconstructing the kinds of problems our Pleistocene ancestors faced, they say. Evolutionary psychologists are on the wrong track. If you want to find out about functions, you should not be looking at T
Abstract
The battle for ‘good science’, so evident in the sociobiology controversy, now continues in the realm of neuroscience. Modelers again clash with hardnosed scientific ‘realists’. From the PANKSEPPs’ point of view, in order to be scientific and ground their own enterprise, evolutionary psychologists ought to pay attention to anatomy and the evolutionary history of the mind.
Moreover, the PANKSEPPs’ criticism of cognitive neuroscience parallels traditional ethological objections to sociobiology, emphasizing not only the ultimate function, but also the development, evolutionary history, and proximate causes of behaviors. This may signal an emerging conflict between evolutionary psychology and ethology. Meanwhile, the PANKSEPPs are in good critical company with many others, such as developmentally oriented scientists, who argue that the modularity in the adult brain does not reflect a preexisting domain specificity, but is rather a product of epigenesis. Indeed, a new interdisciplinary alliance may be currently emerging, as the critics of sociobiology and evolutionary psychology, in their search for a more ‘holistic’ and ‘pluralistic’ paradigm, have recently discovered ethology!
Finally, we should not forget that evolutionary psychology is an evolving research program. Most likely we will see an eventual rapprochement between cognitive and affective neuroscience. A possible mediator could be the interdisciplinary field of nonverbal communication.
Key words
Sociobiology controversy, neuroscience, ethology, evolutionary psychology, development, nonverbal communication, human nature, modularity.
man’ as a whole). I believe that the PANKSEPPs’ stance may be reflecting something more than an opposition between a cognitive and an affective approach to neuroscience. It could be an expression of a deeper conflict between two neighboring fields: evolutionary psychology and ethology. In this commentary I note how their criticism often coincides with an ethological viewpoint and some of the objections that ethologists have typically had to sociobiology. I also find the PANKSEPPs’ views to be surprisingly in tune with those of the critics of sociobiology. Indeed, a new ‘critical syndrome’ appears to be forming in academia, bringing together researchers from different fields with different types of objections to evolutionary psychology and gene-selectionist sociobiology. Finally I discuss the role of the interdisciplinary field of nonverbal communication as a
evolutionary biology, but rather at neuroanatomy.
Lining up ‘seven deadly sins’ of evolutionary psychology, as they do, may sound a bit harsh. (Well, Konrad LORENZ listed eight deadly sins, but that was of a much broader population, ’civilized
possible mediator between cognitive and affective neuroscience, or between an information oriented and an anatomical approach.
The PANKSEPPs are convinced that the best scientific progress in elucidating the mind will be made on the basis of real, physical knowledge rather than through modeling. In the sociobiology controversy, experimentally oriented scientists voiced a similar criticism of the efforts of sociobiologists, who were perceived as constructing models which had little to do with reality. Sometimes the accusation went so far as to allege that sociobiologists actually believed in the reality of such things as average gene frequencies, rather than considering these to be mere calculations. And just in the same way in the earlier (and overlapping) IQ controversy, psychometricians had been accused by largely the same critics of ‘reifying’ a mere computation of test scores into a mysterious entity, general intelligence, g. According to the critics, since this was a mere statistical construct, it ‘could not’ exist in anyone’s head.1 For the critics, real science had to do with research at the molecular level (cf. SEGERSTRÅLE 2000, especially chapters 13 and 14).
The sociobiologists, for their part, felt perfectly justified in constructing models, as did the psychometricians in making their correlations and calculations. For them, these were quite legitimate approaches in their respective fields. They had no doubts, meanwhile, that their results, obtained by a different scientific route, would be compatible with future molecular level research.
Nowhere is the conflict clearer between a modeling and a ‘realist’ approach than in the case of the meaning of the ‘gene’. For instance, molecular biologist Gunther STENT (1977) called DAWKINS’ notion of the selfish gene ‘a heinous terminological sin’, pointing out that the ‘true’ gene was ‘unambiguously’ that unit of genetic material which encoded the amino acid sequence of a particular protein (a cistron)—and, in addition, that it was not selfish! Indeed, laboratory geneticists sometimes seemed especially displeased with sociobiological models sometimes one got the feeling that they wanted sociobiologists to go into the lab and do ‘real’ scientific research on genes (e.g., HOWE/LYNE 1992; SEG-ERSTRÅLE 2000, pp386–387).
In other words, in these controversies there was a clear dividing line in regard to what was ‘good science’. The critics felt remarkably free to accuse sociobiologists of scientific ‘error’ for holding views and using methods that these scientists themselves considered standard in their own field! Some critics went as far as using the ‘errors’ of sociobiologists as evidence that the sociobiological agenda ‘must’ be politically motivated. According to them, the sociobiologists’ scientific quest ‘could not’ be driven by scientific interest, because the only interesting science was done at the molecular level (e.g., LEWONTIN 1975; CHOROVER 1979).
Now in the case of the PANKSEPPs vs. evolutionary psychology we seem to have a similar opposition between two views of ‘good science’: a ‘hard data’ approach conflicting with a model-happy one. Take the PANKSEPPs’ criticism of the computational cognitive science models used by evolutionary psychologists. According to them, real science is done at the anatomical, physical level. PET scans and other fancy new methods for showing brain activity are at best correlational, and can easily be misleading about the true neural processes. They also point out that evolutionary psychologists, following the lead of cognitive scientists, use a digital model of mind. But the mind is analog, they protest.
In other words, the battle for good science, so evident in the sociobiology controversy, now continues in the realm of neuroscience. Modelers again clash with hardnosed scientific ‘realists’. But if the sociobiologists were criticized for telling adaptive ‘just-so’ stories about hypothetical genes ‘for’ behaviors and speculating on the basis of very little evidence, the present authors go one step further. They accuse the evolutionary psychologists of ignoring a whole body of evidence that actually exists: the findings from comparative neuroscience! And the consequences are grave, the PANKSEPPs maintain. Evolutionary psychologists feel free to postulate a system of massive modularity in the neocortex. But from the point of view of comparative neuroanatomy, such modules could not have evolved.
To rub this in, they cite TOOBY and COSMIDES’ (1992) own call for consistency across scientific fields. TOOBY and COSMIDES criticize social scientists for what they call the Standard Social Science Model, which regards the human mind as a generalpurpose machine for learning. Such a model is an evolutionary impossibility, according to them. The PANKSEPPs have no direct argument with this. Rather, they point out that when it comes to their own research, TOOBY and COSMIDES are not applying their own principle: they do not ground themselves on already existing knowledge about the brain.
The whole premise of evolutionary psychology is wrong, say the PANKSEPPs. The problem is not the assumption of modularity per se, but evolutionary psychologists are looking for modules in the wrong place. Any claim about the structure of the human mind/brain would have to take into account its evolutionary history. If routinized behaviors of the ‘module’ kind exist and confer survival value on humans, these are bound to be of a kind we share with
Evolution and Cognition ❘ 7 ❘ 2001, Vol. 7, No. 1
other mammals. This means we had better look for modules in older, subcortical parts of the brain, particularly those parts that deal with emotions, those important motivators for animal survival. (Here they invoke not only Paul MACLEAN’S famous ‘triune brain’ but also E. O. WILSON’s opening statements in Sociobiology). This is why, they argue, comparative, affective neuroscience cannot be ignored.
And they have an additional argument. Evolutionary psychologists have erroneously taken the most recent and particular characteristic of the human brain—our large neocortex—expecting to find modules there. They may believe they have found some modules in the adult brain, but any such modules are likely to be a product of development, they point out. In contrast to the evolutionary psychologists, the PANKSEPPs envision the individual as initially working with a type of general purpose program; it is only later during ontogenesis that more specialized modules are being formed. So for the PANKSEPPs, cognitive modules, to the extent they can be found, are actually by-products, not primary adaptations.
Here we see that the PANKSEPPs adhere to GOULD’s and LEWONTIN’s ‘spandrel’ thinking (GOULD/ LEWONTIN 1979), according to which many traits that sociobiologists see as direct adaptations may actually be mere by-products of evolution, just like some details of architecture—spandrels—are created around vaults. This would seem to put them squarely in the same camp as the ‘anti-adaptationist’ critics of sociobiology. And they in fact go a step further, arguing, with GOULD (1991), that human language, too, is likely to be an evolutionary byproduct.2 And if language is a mere spandrel, then focusing on language as the model module is obviously misdirected. (This latter point is an unnecessary piece of overkill, in my view, because the PANK-SEPPs’ argument does not hinge on the nature of language. True, if it were possible to conclusively prove that language was a mere spandrel, they would have a point. But GOULD is operating rather on the level of principle, while for many following CHOMSKY’s and PINKER’s work, the idea of a particular language module does appear persuasive. Might it not be possible to regard language as a module without taking the next step with PINKER (1994, 1997a) and argue from the modularity of language to a view of massive modularity?).
From the PANKSEPPs’ point of view, then, in order to be scientific and ground their own enterprise, evolutionary psychologists ‘ought to’ pay attention to anatomy and the evolutionary history of the mind. They are dismayed that COSMIDES and TOOBY in a recent contribution went so far as to attempt to deal with emotion without making any mention of affective neuroscience (COSMIDES/TOOBY 2000). What could be some reasons for this neglect among the leading theorists in evolutionary psychology?
In the first place, evolutionary psychologists have their hands full. They are busy fighting a multi-front battle: against cultural anthropologists and mainstream psychologists on the one hand, and against sniping critics of ‘adaptationism’ on the other. In many regards, they have inherited the mantle of sociobiology, and with this, its critics.3 Despite their attempt to emphasize universal features of humans and ‘the psychic unity of mankind’, and thus trying to fend off accusations of racism and biological determinism, they are labelled ‘ultra-DARWINIANS’ by GOULD and other critics (GOULD 1997a–b; ELDREDGE 1995; ROSE 1998). They have to create legitimacy for their field at the same time as they have to establish a viable research program.
Predictably, too, the reception of evolutionary psychology among mainstream psychologists has not been too hospitable. But there were potential allies among scholars in other fields which also deal with the mind, and whose approach is similar or at least compatible with the methods typically used in evolutionary psychology. Cognitive science seemed the natural way to go. By associating itself with cognitive science, evolutionary psychology could ride on the wave of the triumphant cognitive revolution. It could be linked to booming research fields such as artificial intelligence, and to novel methods for monitoring cognitive processes. Computational algorithms would provide the necessary theoretical rigor, while new imaging techniques would provide the physical correlates for their claims.
The PANKSEPPs are duly respectful of the cognitive revolution—it is just that they feel that the next revolution, the neuroscientific one, did not have a chance to make its mark before the surge of interest in evolutionary explanation. The new kin selection paradigm swept people off their feet, they complain. And here it becomes clear that, unlike sociobiologists and evolutionary psychologists, the PANK-SEPPs are interested not only in ultimate answers about function, but also in the development, evolutionary history, and proximate causes of behaviors. In other words, they want to emphasize the importance of all of TINBERGEN’s famous ‘four questions’ (TINBERGEN 1963). As we see, they are particularly concerned about developmental aspects.
Interestingly, a very similar complaint about evolutionary explanations ‘taking over’ can be found among bona fide ethologists in their comments on the 1970’s sociobiological revolution. Just like the present authors, many ethologists felt that in the 1970s their field, too, was on the brink of making interesting discoveries and solidifying a new, complex, paradigm for the study of animal behavior (as demonstrated, e.g., by BATESON/HINDE 1976). But when the exploitable-seeming new sociobiological framework arrived, many ethologists chose to focus on only one of TINBERGEN’s four questions, the question about function, and this served to drive ethology into a narrow direction (BATESON/KLOPFER 1989). (Recently, though, the sociobiology ‘gold rush’ appears to be over, and the tide has turned back toward a more complex ethological approach, e.g., KREBS/DAVIES 1997).
But the PANKSEPPs do not only criticize evolutionary psychology on scientific grounds, they also warn the field about potential political consequences. Here it is not clear exactly what they have in mind. The political critics of sociobiology actually reasoned in a number of different ways about sociobiology. The idea of a genetically based universal human nature was anathema for them, because they saw this as connected to genetic determinism and other evils. But a further belief was that ‘bad science’ (for the critics, sociobiology and IQ research) would inevitably lead to undesirable social consequences. As I have argued (SEGERSTRÅLE 2000), there is no obvious reason why ‘bad’ science (however defined) would be particularly prone to social abuse; it seems to me that ‘good’ science in the wrong hands would actually be more dangerous. If we apply this to evolutionary psychology, there would seem to be no inherent connection between, say, belief in modularity of mind and risk of political abuse. The PANKSEPPs may simply mean that, just like sociobiology, evolutionary psychology is going to be politically attacked as a deterministic, dangerous doctrine about human nature.
If that is what they mean, that has already happened. For instance THORNHILL and PALMER’s (2000) recent book about rape created quite a stir (e.g., COYNE 2000; SHULEVITZ 2000; for a response see TOOBY/COSMIDES 2000). And recently there was a collection of critical essays on evolutionary psychology (ROSE/ROSE 2000), which was surprisingly well received in the popular press at least in the United Kingdom. I take this to mean that there is a blossoming market for this kind of moral–cum–scientific criticism both in the UK and the US—just as there is a great market for books that explain why we do what we do. Indeed, a certain symbiosis appears to prevail between the writers of popular evolutionary psychology books and their critics—a phenomenon observed already in the sociobiology controversy (SEGERSTRÅLE 2000).
Of course, evolutionary psychologists have made the situation rather difficult for political critics. Well aware of the attacks on sociobiology, they have carefully stated that they are interested in the evolved human mind, not genes, and in a universal human nature, not human differences. They do not employ population-genetic models, and do not address genetic differences between populations.4 They explicitly distance themselves from human behavioral genetics (something WILSON needed for his sociobiological approach). This makes it hard to construe evolutionary psychologists as racists—the standard move in left-wing criticism of biological theories of human nature, and one that was much employed in the sociobiology controversy (cf. SEG-ERSTRÅLE 2000, esp. ch. 2, 9, and 15). Finally, they are uninterested in IQ differences, which means they are not connectable either to the various standard accusations against psychometricians (cf. SEGER-STRÅLE 2000, esp. ch. 10, 12, 13, 14). Indeed, a recent primer in evolutionary psychology sounds remarkably ‘politically correct,’ repudiating explicitly both the notion of race and the idea of ‘general intelligence’ (COSMIDES/TOOBY 1997).
Whatever the evolutionary psychologists say, however, it is clear that the former critics of sociobiology do not like evolutionary psychology, as little as they like sociobiology (and they typically criticize these fields together). In their search for scientific and moral/political counter-arguments the opponents to evolutionary psychology have generated a plethora of different criticisms, not all necessarily compatible with one another. This is déjà vu, and one needs only look back at the sociobiology controversy. Still, the tone is now more muted, and the critique more scientific and not as personal and nasty. One reason for this more cautious approach may be that it has become harder to stir up general outrage around claims about the evolutionary basis of human behavior. At the turn of the millennium the idea of genes influencing behavior has become more acceptable to the general public. In fact, some worry that the situation has totally flip-flopped. They fear a new doctrine of ‘genetic essentialism’ will be holding genes increasingly responsible for our behavior (NELKIN/LINDEE 1995; cf. SEGERSTRÅLE 2000, ch. 19).
This is why veteran critic of sociobiology, Steven ROSE (2000), coauthor of Not In Our Genes (LEWONTIN/ROSE/KAMIN 1984), does not any longer
Evolution and Cognition ❘ 9 ❘ 2001, Vol. 7, No. 1
declare that nothing is in our genes. In his criticism of evolutionary psychology he now leans on a battery of other arguments, particularly well-known anti-adaptationist ones. What is interesting, however, is the remarkable convergence between the criticism coming from this long-standing critic of sociobiology (whom I have called ‘Britain’s LEWONTIN’) and the PANKSEPPs’ present critique of evolutionary psychology. One reason for this could be that ROSE is, after all, a neurospsychologist with a ‘realist’ bent. Still, in both ROSE and the PANKSEPPs, I see something like a cluster of similar critical ideas. Just like the PANKSEPPs, ROSE criticizes the evolutionary psychologists’ conception of the brain as an information processing machine. Brains/minds do not only deal with information, ROSE points out: they deal with meaning. And meanings involve emotions! ROSE goes on to question the idea that modules have evolved ‘quasi independently’ and that they are primary, that is, that they underlie proximal mechanisms such as motivation. Finally, ROSE points to the survival value of emotion and invokes recent research in ‘the emotional brain’ by people such as Antonio DAMASIO and Joseph LEDOUX.
What I perceive here is an emerging alliance between ethologists and left-wing critics of sociobiology and evolutionary psychology. It seems that the critics of sociobiology, in their resistance to geneselectionism, have grasped for a more ‘holistic’ and ‘pluralistic’ paradigm—and voilà, they have found ethology! This means that the critics of sociobiology and evolutionary psychology have come a long way from their initial position, which seemed to dismiss even the thought of an evolutionary basis for human behavior (cf. for instance, ALLEN et al. 1975). And look what more we find! Another element in the evolving new moral–cum–scientific vision of the new opposition appears to be a belief in group selection, that heretic view so irritating to adherents of gene selectionism (cf. SEGERSTRÅLE 2000, pp383– 384). This belief has certainly been stimulated by the new study of SOBER/WILSON (1998). So, here we now have ROSE, always trendy, topping off his critique of evolutionary psychology with an argument for group-selection. The PANKSEPPs, too, seem to like group selection, a way of thinking clearly compatible with a (LORENZIAN) ethological model.
But criticism of evolutionary psychology does not only come from the earlier critics of sociobiology. It comes from a number of directions: developmentally oriented biologists and psychologists (e.g., BATESON/MARTIN 2000), biological systems theorists (e.g., OYAMA 2000), a variety of researchers and theorists of cognitive processes (e.g., VAN GELDER/PORT 1995; HUTCHINS 1996), and, finally, even one type of cognitive scientists: students of developmental cognitive neuroscience (e.g., JOHNSON 1997). These critics typically argue that the computational model of the mind does not cover many important aspects of the mind and living systems, such as internal regulation, coordination, and feed-back mechanisms, or they protest that human intelligence is of a complex type that simply cannot be translated into machine language.
Not surprisingly, researchers on development have found the model used by evolutionary psychology too deterministic. Among other things they point out that many supposedly ‘innate’ competencies actually presuppose learning during an organism’s lifetime (e.g., BATESON 1987; BATESON/ MARTIN 2000). Still, it seems to me that they may be sometimes caricaturing the evolutionary psychologists’ position in order to drive home their point. It often seems unclear just how their own views differ, particularly since TOOBY and COSMIDES already in 1992 emphasized that they do not expect modules to be ‘hardwired’ but only to develop reliably. The question is what this means. Those who study development may want greater explicit recognition of the variety of phenotypes that can develop depending on environmental conditions, and of the de facto unpredictability of animal and human behavior (see SEGERSTRÅLE 2000, pp378–379)—or might they perhaps wish to suggest that organisms ‘choose’ their environments?
An interesting criticism comes from a developmental psychologist, Annette KARMILOFF-SMITH, who challenges one of the arguments used to boost nativism and modularity: the so-called ‘double dissociation’ found in WILLIAMS syndrome patients. People with this syndrome are said to be good at recognizing faces, language, and social interaction but have difficulty with spatial cognition, numbers, and problem-solving. KARMILOFF-SMITH studied more closely children and adults with this developmental disorder, and concluded that the assumption that WILLIAMS syndrome patients were otherwise normal was incorrect. In fact, WILLIAMS syndrome patients differed from controls in their dealing both with faces and with language and grammar. In regard to faces, the children instead of adopting holistic, configurational, approach put together faces feature–by–feature. This she interpreted to mean that they in ontogeny had developed a novel, compensatory strategy for dealing with information. They also followed an unusual pathway
Evolution and Cognition ❘ 10 ❘ 2001, Vol. 7, No. 1
in their language acquisition. According to her, this was evidence for the dynamic development of the brain and the interaction of its different parts in ontogeny (KARMILOFF-SMITH 2000, 1998, 1992).
And there is more evidence for flexibility in the developing brain. For instance, brain imaging findings of face processing (in normal infants) show increased specialization and localization of face-processing circuits during the first year of life. Language, too, shows progressive specialization of this sort: it is processed in both hemispheres until about the age of six when it (for right-handed people) goes to the left hemisphere (KARMILOFF-SMITH 2000; JOHNSON 1997).
These kinds of findings would seem to support the PANKSEPPs’ point that the modularity in the adult brain does not reflect a pre-existing domain specificity of the brain, but is rather a product of epigenesis. Although the PANKSEPPs do not elaborate on this, one would assume that they would concur with the distinction between two views of development: a ‘regulatory’ vision as opposed to a ‘mosaic’ one. While the ‘mosaic’ kind is represented by the evolutionary psychologists’ vision of ‘the mind as a Swiss army knife’ and presupposes tight genetic control on epigenesis, the regulatory model involves a probabilistic epigenesis under broad genetic control, but flexible.5 KARMILOFF-SMITH specifically suggests that there exists a number of different evolved learning mechanisms which during ontogeny may “each discover inputs from the environment that are more or less suited to their form of processing”. This is why she calls for a closer study of the Swiss army knife’s ontogenesis (KARMILOFF-SMITH 2000).
More alarming, perhaps, for evolutionary psychologists—and giving an indirect boost to the PANKSEPPs—is the recent removal of one of the cornerstones of the whole modular theory. The very father of modularity, Jerry FODOR himself (FODOR 1983), appears to have had a change of mind (pun intended!), and now seems devoted to attacking those who describe the mind as a collection of domain-specific special processing machines. This is instantiated by his new The Mind Does Not Work That Way (Fodor 2000), a direct critique of Steve PINKER’s (1997a) How The Mind Works.
Finally, the PANKSEPPs get unexpected support from someone quite sympathetic to sociobiology and a fierce critic of its critics, anthropologist Melvin KONNER. On the face of it KONNER ‘should’ be enthusiastic about evolutionary psychology, but he takes a surprisingly critical stance of PINKER’s 1997 book (KONNER 1998). One of his main complaints is
that PINKER and evolutionary psychologists in their modeling totally ignore the actual anatomical structure of the brain. One cannot make the assumption of homogenous modularity throughout the brain.
In other words, the PANKSEPPs are in good critical company.
But our authors are not intending only to be critical. In fact, they say they wish to help evolutionary psychology. What are, in fact, the possibilities of bringing cognitive and affective neuroscience together? Is there a middle ground that might be reached? In light of the criticism, it would seem prudent for evolutionary psychologists to postulate a limited number of basic modules rather than massive modularity. Plausible candidates for more or less hardwired specialized programs come from animal studies and research on neonates in humans. I believe there is enough evidence, for instance, to speak of a ‘face recognition module’, based on infant research and the demonstrated existence of ‘face neurons’ in monkeys and sheep. Moreover, imitation may be another such basic capability, as may the set of basic emotional expressions, as demonstrated by Paul EKMAN and his associates (e.g., EK-MAN/KELTNER 1997). One indication here is that we have all necessary facial muscles fully developed at birth, another that neonates—even anencephalic ones—react in predictable ways to sweet and sour substances (SEGERSTRÅLE/MOLNAR 1997, pp8–9). Interestingly, even Gerald EDELMAN, otherwise committed to a neural network, ‘constructivist’ view of the mind, believes that there do, in fact, exist a few basic inborn capabilities in humans: among these face recognition, and fundamental affect (SEGER-STRÅLE/MOLNAR 1997, p16).
Many of our basic abilities do appear to be connected to the human face and be of a nonverbal nature. It would seem that the face has important inbuilt capabilities that will later form the basis for the infant’s further emotional development and interactional skills (SEGERSTRÅLE/MOLNAR 1997, p16, see also Part III on ontogeny). Thus, one more candidate for relatively hard-wired equipment may be certain fast-moving ‘facial affect programs’ that enable us to competently send and receive nonverbal messages, identified by Swedish psychologist Ulf DIMBERG (DIMBERG 1997). In other words, nonverbal communication is an important means whereby ‘nature’ is transformed to ‘culture’. Also, recent research on language emphasizes the importance of nonverbal communication as a preparatory stage in language acquisition (VELICHKOVSKY/RUMBAUGH 1996).
Evolution and Cognition ❘ 11 ❘ 2001, Vol. 7, No. 1
Ullica Segerstråle
The PANKSEPPs, with MACLEAN and his triune brain, and WILSON (in Sociobiology) see emotions as primary, because of the evolutionary history of the brain. Still, nonverbal capabilities appear to be at the same time emotional and cognitive. And this may eventually help open the door for a synthesis between cognitive and affective neuroscience.
In general, then, why not relax the modularity idea, concentrate on a few indisputable ones, and postulate different degrees of hardwiredness for different human capabilities based on the best available empirical studies across a variety of fields, while leaving space also for general, undifferentiated learning programs? Also, why not keep the question open as to where exactly some of these circuits are located and how big these modules are. (Perhaps even MACLEAN’s various brains can be seen as modules?) The important thing would seem to be to be able to pool ideas from a variety of fields: cognitive and affective neuroscience, and a wide spectrum of interdisciplinary nonverbal research. That kind of approach would seem to go toward TOOBY and COSMIDES–style consistency across scientific fields, ‘consilience’ of the William WHEWELL type (simultaneous support for an idea from many fields of science) and may be even E. O. WILSON type ‘consilience’ (unity of science) (WILSON 1998).
Unfortunately there is a real obstacle to mutually beneficial cooperation between scientists from different disciplines. Interdisciplinary efforts may give them no credit—on the contrary, they may be ostracized by their ‘home’ discipline, particularly if they are regarded as breaking certain long-standing taboos.
But perhaps affective neuroscience needs to rethink its criticism somewhat. Whatever modules ‘really’ are, and how many ‘really’ exist, the basic theoretical contributions of the evolutionary psychologists are interesting and
Notes
- 1 While they sharply resisted this kind of research in cognitive abilities, the critics expressed interest in finding out about cognitive traits.
- 2 On this topic in 1997 GOULD (1997c) got himself involved in a furious dispute with Steven PINKER (1997b) on the pages of The New York Review of Books.
- 3 WILSON thinks evolutionary psychology is “the same” as sociobiology (personal communication), and so does DAWKINS.
can in principle open up new fruitful avenues of research in other fields—including affective neuroscience. This kind of importation into one field of ideas from another is a well-known device for scientific growth—after all, it was physicists rather than geneticists that created the new field of molecular biology. What the possibilities are for a field to stimulate another at a particular time is dependent on the availability of individuals and their visions and ambitions. Fields develop according to concrete opportunities perceived and particular initiatives taken—not according to how they ‘ought to’ develop from some abstract scientific point of view.
Finally, we should not forget that evolutionary psychology is an evolving research program. There are already signs that the evolutionary psychologists are backing off somewhat from their initial militancy. For instance, in COSMIDES and TOOBY’s recent primer the assumption of the Environment of Evolutionary Adaptation seems to be relaxed, the assumption of modularity looks less massive, and epigenesis is more explicitly factored in. Emotions, too, are increasingly brought in, a potential harbinger of the next step of the evolving program of evolutionary psychology. If this kind of rapprochement between the evolutionary psychologists and their critics is taking place, it is only something that is typical of science. Indeed, in the future, we may expect both (all?) sides to integrate some of their opponents’ arguments into their own explanatory frameworks—without necessarily giving their critics explicit credit. We may even see a convergence between seemingly totally opposed positions (remember the DARWINISTS and MENDELIANS
Author’s address
Ullica Segerstråle, Department of Social Sciences, Illinois Institute of Technology, Suite 116, Siegel Hall, 3301 South Dearborn Street, Chicago IL 60616, USA. Email: segerstrale@iit.edu
fighting each other only to be united in the Modern Synthesis?). But if and when this happens, we will probably not have a synthesis with big fanfare. Academia being what it is, both sides are likely to claim victory and get out.
- 4 Something that WILSON was challenged into doing by critics who said that he did not address human variation.
- 5 Note, however, that although her arguments supports that of our authors, she is actually citing data from imaging processes which the PANKSEPPs may consider unreliable!
- 6 KARMILOFF-SMITH herself believes a regulatory approach is more likely to reach the complexity required by cortical functions. She suggests, however, that a deterministic view of development could be applicable to other parts of the brain.
Evolution and Cognition ❘ 12 ❘ 2001, Vol. 7, No. 1
References
- Bateson, P. P. G. (1987) Biological approaches to the study of behavioral development. International Journal of Behavioral Development 10: 1–22.
- Bateson, P. P. G./Klopfer, P. H. (1989) Preface. In: Bateson, P. P. G./Klopfer, P. H. (eds) Whither Ethology? Perspectives in Ethology 8. Plenum Press: New York, London, pp. v–viii.
- Bateson, P. P. G./Martin, P. (2000) Design for A Life: How Biology and Psychology Shape Human Behavior. Touchstone Books: New York.
- Chorover, S. (1979) From Genesis to Genocide. MIT Press: Cambridge MA.
- Cosmides, L./Tooby, J. (1992) The psychological foundations of culture. In: Barkow, J/Cosmides, L./Tooby, J. (eds) The Adapted Mind. Oxford University Press: New York.
- Cosmides, L/Tooby, J. (1997) Evolutionary Psychology: A Primer. Retrieved on 15 Feb 2001 from the World Wide Web: http://www.psych.ucsb.edu/research/cep/primer.html
- Cosmides, L/Tooby, J. (2000) Evolutionary psychology and the emotions. In: Lewis, M./Haviland, J. (eds) The Handbook of Emotions. 2nd edition. Guilford: New York, pp. 91–116.
- Coyne, J. (2000) Of vice and men. Review of “A Natural History of Rape” by R. Thornhill and C. Palmer. New Republic 04.03.00: 27–34.
- Dimberg, U. (1997) Psychophysiological reactions to facial expressions. In: Segerstråle, U./Molnar, P. (eds) Nonverbal Communication: Where Nature Meets Culture. Lawrence Erlbaum: Mahwah NJ, pp. 47–60.
- Ekman, P./Keltner, D. (1997) Universal facial expressions of emotions: An old controversy and new findings. In: Segerstråle, U./Molnar, P. (eds) Nonverbal Communication: Where Nature Meets Culture. Lawrence Erlbaum: Mahwah NJ, pp. 27–46.
- Eldredge, N. (1995) Reinventing Darwin. The Great Debate at the High Table. John Wiley and Sons: New York.
- Fodor, J. (1983) The Modularity of Mind. MIT Press: Cambridge MA.
- Fodor, J. (2000) The Mind Doesn’t Work That Way. MIT Press: Cambridge MA.
- van Gelder, T./Port, R. F. (ed) (1995) Mind as Motion: Explorations in the Dynamics of Cognition. MIT Press: Cambridge MA.
- Gould, S. J. (1991) Exaptation: A crucial tool for evolutionary psychology. Journal of Social Issues 47: 43–65.
- Gould, S. J. (1997a) Darwinian fundamentalism. The New York Review of Books, June 12: 34–37.
- Gould, S. J. (1997b) Evolution: the pleasures of pluralism. The New York Review of Books, June 26: 47–52.
Gould, S. J. (1997c) Evolutionary psychology: An exchange. New York Review of Books, October 9.
- Gould, S. J./Lewontin, R. C. (1979) The Spandrels of San Marco and the Panglossian Paradigm: A Critique of the Adaptationist Programme. Proceedings of the Royal Society of London B 205: 581–598.
- Howe, H./Lyne, J. (1992) Gene Talk in Sociobiology. Social Epistemology 6: 109–63.
- Hutchins, E. (1996) Cognition in the Wild. MIT Press: Cambridge MA.
- Johnson, M. (1997) Developmental Cognitive Neuroscience: An Introduction. Blackwell: Oxford.
- Karmiloff-Smith, A. (1992) Beyond Modularity: A Developmental Perspective. MIT Press: Cambridge MA.
- Karmiloff-Smith, A. (1998) Development itself is the key to
understanding developmental disorders. Trends in Cognitive Science 2: 389–398.
- Karmiloff-Smith, A. (2000) Why babies’ brains are not Swiss army knives. In: Rose, H./Rose, S. (eds) Alas, Poor Darwin: Arguments Against Evolutionary Psychology. Jonathan Cape: London, pp. 144–156.
- Konner, M. (1998) A piece of your mind. Review of “How the Mind Works” by S. Pinker. Science 281: 653–654.
- Krebs, J. R./Davies, N. B. (1997) Behavioural Ecology. Fourth Edition. Blackwell Scientific Publications: Oxford.
- Lewontin, R. C. (1975a) Genetic aspects of intelligence. Annual Review of Genetics 9: 387–405.
- MacLean, P. D. (1990) The Triune Brain in Evolution. Plenum Press: New York.
- Nelkin, D./Lindee, M. S. (1995) The DNA Mystique. Freeman: New York.
- Oyama, S. (2000) The Ontogeny of Information: Developmental Systems and Evolution. Duke University Press: Durham NC.
- Pinker, S. (1994) The Language Instinct. HarperCollins: New York.
- Pinker, S. (1997a) How the Mind Works. W. W. Norton: New York.
- Pinker, S. (1997b) Evolutionary psychology: An exchange. New York Review of Books, October 9.
- Rose, S. (1998) Lifelines. Oxford University Press: New York.
- Rose, S. (2000) Escaping evolutionary psychology. In: Rose, H./Rose, S. (eds) Alas, Poor Darwin: Arguments Against Evolutionary Psychology. Jonathan Cape: London, pp. 247–265.
- Rose, H./Rose, S. (eds) (2000) Alas, Poor Darwin: Arguments Against Evolutionary Psychology. Jonathan Cape: London.
- Segerstråle, U. (2000) Defenders of the Truth: The Battle for Science in the Sociobiology Debate and Beyond. Oxford University Press: Oxford.
- Segerstråle, U./Molnar, P. (eds) (1997) Nonverbal Communication: Where Nature Meets Culture. Lawrence Erlbaum: Mahwah NJ.
- Shulevitz, J. (2000) Evolutionary psychology teaches Rape 101. Dialogues. Slate (online magazine), January 13. Retrieved on 15 Feb 2001 from the World Wide Web: http:// slate.msn.com/code/Culturebox/
Culturebox.asp?Show=1/13/2000&idMessage=4368
- Sober, E./Wilson, D. S. (1998) Unto Others: The Evolution and Psychology of Unselfish Behavior. Harvard University Press: Cambridge MA.
- Stent, G. S. (1977) You can take ethics out of altruism but you can’t take the altruism out of ethics. Hastings Center Report 7: 33–36.
- Thornhill, R./Palmer, C. (2000) A Natural History of Rape. MIT Press: Cambridge MA.
- Tinbergen, N. (1963) On aims and methods of ethology. Zeitschrift für Tierpsychologie 20: 410–433.
- Tooby, J./Cosmides, L. (2000) Letter to the editor of New Republic (response to Coyne). Retrieved on 15 Feb 2001 from the World Wide Web: http://www.psych.ucsb.edu/research/cep/
- Velichkovsky, B./Rumbaugh, D. (eds) (1996) Communicating Meaning: The Evolution and Development of Language. Lawrence Erlbaum: Mahwah NJ.
- Wilson, E. O. (1975) Sociobiology: The New Synthesis. Harvard University Press: Cambridge MA.
- Wilson, E. O. (1998) Consilience: The Unity of Knowledge. Alfred Knopf: New York.
Evolution and Cognition ❘ 13 ❘ 2001, Vol. 7, No. 1
“La plus ça change…”
Response to a Critique of Evolutionary Psychology
ANKSEPP & PANKSEPP are not alone in their criticisms of evolutionary psychology. In fact, controversies sparked by this ‘new’ discipline abound and have even merited front page coverage in a recent issue of the NY Times’ ‘Science Times’ (GOODE 2000). It is clear, however, that most of the critics, including P&P, are against neither evolution nor psychology—nor even the use of evolutionary approaches in psychology. Indeed many, like P&P, are themselves practitioners of what might be called evolutionary psychology without the capital E capital P P
Abstract
PANKSEPP & PANKSEPP are not alone in their criticisms of Evolutionary Psychology. It is clear, however, that most of the critics, including P&P, are against neither evolution nor psychology—nor even the use of evolutionary approaches in psychology. Criticism has, rather, focused on a particular approach taken by a very small but highly visible group of individuals who share a specific stance on a set of related issues. I share P&P’s concerns about the rigidity and doctrinal restrictiveness of “some currently fashionable versions of evolutionary psychology”. But this debate is not new, and I believe that as with most controversies in science, its sparks have propelled us more steps forward than back.
Key words
Cognition, comparative method, evolutionary psychology, modularity, philosophy of science, science studies.
(see e.g., MEALEY 1994, in press; MILLER 2000a, 2000b; THIESSEN 1998; WILSON 1994). Criticism has, rather, focused on a particular approach taken by a very small but highly visible group of individuals who share a specific stance on a set of related issues. In their target paper, P&P variously refer to this subset of theorists and their positions, respectively, as “a new breed of evolutionary psychologists” (p3) and “the most visible form of evolutionary psychology” (p30). Speaking as one who would prefer to see multidisciplinary integration rather than interdisciplinary derogation, I share P&P’s concerns about the rigidity and doctrinal restrictiveness of “some currently fashionable versions of evolutionary psychology” (p6). But this debate is not new, and I believe that as with most controversies in science, its sparks have propelled us more steps forward than back.
The Seven Sins
The environment of evolutionary adaptedness (EEA)
John TOOBY, Leda COSMIDES, David BUSS, Randy THORNHILL, and others, describe evolutionary psychology as a kind of ‘reverse engineering’ through which we try to decipher the components and workings of the human brain/mind by analyzing the types of ‘problems’ it has evolved to ‘solve’ in its ‘environment of evolutionary adaptation’ (e.g., BUSS 1995; COSMIDES/TOOBY 1997; THORNHILL 1997; TOOBY
1999; TOOBY/COSMIDES 1990, 1992). Of course, the human brain/mind has been evolving ever since there was first a neuron, and the problems that it has evolved to solve have waxed and waned and reversed and conflicted and interacted in countless ways over that immense span of time. The EEA of the human brain/mind is thus, not a single definitive environment that we might reconstruct as we might construct a museum diorama, but rather, a multidimensional hyperspace of the net vectors of every selection pressure that ever impinged upon our ancestors (DALY/WILSON 1999; MEALEY 2000).
Stated as such, the EEA is a potent concept. Unfortunately, it is a concept that has been trivialized and reduced to a much more simplistic but manageable notion, i.e., ‘the Pleistocene’, which, as a heuristic for making novel hypotheses about the brain/mind, is virtually useless. This common substitution of a
Evolution and Cognition ❘ 14 ❘ 2001, Vol. 7, No. 1
simple concept for a more complex one needs to be pointed out—much as does the reification of the term ‘gene’ in the commonly used phrase ‘a gene for x behavior’—but however unfortunate such substitution may be, it is a ubiquitous phenomenon in the social process that is the communication of science and spread of knowledge (see also YOUNG/PERSELL 2000). The good evolutionary psychologist might need to be more careful, but need not be damned for this sin.
Species-centrism
Lack of a true comparative foundation is, indeed, a weakness of a modern evolutionary psychology that has forgotten the word ‘ethology’ and the name ‘TINBERGEN’ (see DALY/WILSON 1999). Although comparative psychology and ethology were antagonistic disciplines through much of their early history (JAYNES 1969), most of their differences were resolved with the development and joint use of new techniques in neuroscience and neuroendocrinology (HINDE 1966); even more recently, progress in computer modeling and evolutionary taxonomy have made comparative analyses both easier and more valuable (HALL 1994; HARVEY/PURVIS 1991; LARSON/LOSOS 1996). Still, comparative psychology and human ethology seem to be undergoing major decline—or, if not decline, at least further isolation from what is now mainstream psychology: conferences and journals which once attracted and even showcased research on both human and other species now tend to specialize in one or the other. (See discussions in HIRSCH 1987 and MEALEY 2001.)
A case in point to illustrate the consequences of this neglect can be drawn from a highly visible research area: language. According to evolutionary psychologists, language is the prime exemplar of a recently-evolved, complex, but uniquely human faculty (PINKER 1994, 1997a, 1997b). Unfortunately, to many this status implies that comparative analysis is useless. Indeed, after the once-popular ape studies documented the species-specific nature of human language, comparative studies virtually disappeared. Yet the notion that our language capacity has its basis in a set of modular functions is one that absolutely begs for comparative analysis. For example, for decades psycholinguists cited the phenomenon of ‘categorical perception’ of human speech sounds as evidence for the specialness of human language (EIMAS 1985; EIMAS/MILLER 1992; WERKER/LALONDE 1988; LENNEBERG 1967; LIBERMAN/ MATTINGLY 1989; ZATORRE et al. 1992) without bothering to determine whether that particular component of language was indeed, human-specific. Comparative studies, however, show that it is not (HAUSER 1996, 2000); categorical perception is, rather, a preadaption for language that has its basis in some other feature or function of the brain. Of course, this insight does not negate the argument that human language is special and requires study of human subjects—but it does highlight P&P’s argument that evolutionary psychologists, by being too speciescentric, have ignored a tremendous resource for hypothesis-generation and hypothesis-testing, as well as a huge and valuable extant literature (e.g., HAUSER 1996, LIEBERMAN 1977).
Adaptationism
The possibility that language emerged from a set of pre-adaptations, or that language itself is a ‘spandrel’ (GOULD 1991; GOULD/LEWONTIN 1979) has been hotly debated (PINKER/BLOOM 1990). I happen to think that language is not a spandrel. But I also happen to think that others of our most complex attributes might be: specifically, our intelligence and our capacity for self-reflection, which I believe are likely to be spandrels of our Theory of Mind.
My own opinion about what is or isn’t a spandrel is not particularly relevant here, but the idea that very complex seemingly designed functions and processes might be spandrels is, as P&P claim, an idea that is not only disregarded by most evolutionary psychologists, but unfairly attacked and even mocked (e.g., ALCOCK 1998). If evolved features of the human brain/mind are likely to be modular as P&P’s brand of evolutionary psychologist claims, then indeed, the most likely place for us to search for spandrels is amongst those features that seem the most generalized and un-modular… such as the ‘g– factor’ of intelligence and the ‘binding factor’ or ‘seamless nature’ of self-awareness.
Massive modularity
Since the structure of the mind is, in some sense, dependent upon the structure of the brain that houses it, it might be the case that some functional mechanisms of the mind map directly onto certain structures of the brain. While such direct mapping is not a prerequisite for psychic modularity (FODOR 1983), it is plausible (SHALLICE 1988, 1991), and the fact that different parts of the brain do exhibit functional specialization has been known for over a hundred years. Not only are specialized structures
Evolution and Cognition ❘ 15 ❘ 2001, Vol. 7, No. 1
and neural circuits known to regulate such complex operations as sleep and visual–motor coordination, but the agreement of functional maps of the brain with cytoarchitectural maps and projection maps is ‘stunning’ (KOLB/WISHAW 1990) and ‘precise’ (GAZ-ZANIGA 1989). Together with evidence from comparative anatomy (e.g., PANKSEPP 1992, 1998), this suggests an evolutionary pattern of cumulative layering of new brain structures (MACLEAN 1990), rather than a single ‘general purpose’ structure.
P&P not only acknowledge, but have helped to document the existence of, extensive brain modules; their beef is not with the concept of modules, but with the unquestioned assumption of modules—especially at the cortical level, where evidence is sparse (FINLAY/DARLINGTON 1995). They further note that evolutionary psychology’s characterisation of an adaptation as compared to an exaptation or a spandrel (‘complexity, economy, efficiency, reliability, precision and functionality’: BUSS et al. 1998) can be equally applied to various non-modular, and even domain-general, processes. Modern discoveries in neuroscience—such as RAMACHANDRAN’s dramatic demonstrations of cortical rewiring of perceptual maps (RAMACHANDRAN/BLAKESLEE 1998)—raise questions about the fixity, and therefore, the supposed rigid prescription of what otherwise appear to be modular faculties.
I suspect that the delineation of ‘modular’ processes and organs from ‘non-modular’ processes and organs will never be so neat as the picture that FODOR (1983) originally laid out—even FODOR thinks that evolutionary psychologists have carried the concept too far (FODOR 2000). More likely, whether one chooses to admit a particular functional neurological pathway to the club of ‘modules’ or prefers to consider it an exaptation, a spandrel, a “fixed action pattern” (SCHLEIDT 1974), “prepared learning” (SE-LIGMAN 1970), or a case of “tinkering” (JACOB 1977) will turn out to be more a matter of personal opinion than neuropsychology. This too, is not a new debate, and the answer is not going to be of the either/or form.
Conflation of emotions and cognitions
The relationship between emotions and cognitions has also been an ongoing subject of debate. Phenomenologically, does one arise first and then trigger the other? Or are they simultaneous? If simultaneous, are they two aspects of the same thing? If so, then why do we experience them as different? Or do we? Or when do we? If different, is each thought or emotion discrete? Or do they blend? And of course, what is the underlying neurophysiology? (See DAMASIO 1994, 1999; EKMAN/DAVIDSON 1994; GRIFFITHS 1990, 1997; LAZARUS 1991; NESSE 1991, 1992a, 1992b; PANK-SEPP 1998; PLUTCHIK 1980, 1992; ROLLS 2000.)
Emotions and cognitions both serve to motivate the organism: they promote action. Indeed, the words ‘emotion’ and ‘motivation’ share the same Latin root. If emotions and cognitions share function, then it is most likely that they are designed to be experienced together—and of course, it is easy to conflate them if/when we experience them together. One of my favorite quotes on this topic comes from PLUTCHIK (1980); he suggests that:
“The whole cognitive process evolved over millions of years in order to make the evaluations of stimulus events more correct and the predictions more precise so that the emotional behavior that finally resulted would be adaptively related to the stimulus events. It is in this sense that cognitions are in the service of emotions.” (p303)
So emotions are generated first, which are honed and articulated by cognitions, and then expressed as behavior… Or do cognitions first filter and assess stimuli, then direct them to the proper emotion module for action!? No matter which came first phylogenetically, the inputs to various brain/mind structures may be directional or reciprocal, simultaneous or sequential. Which parts of this clearly joint phenomenon are labelled ‘emotion’ and which ‘cognition’ depends in large degree on the breadth of one’s sweep and the differential jargon of disciplines (MALLON/STICH 2000). We know better than to speak of nature versus nurture, and we know better than to speak of learning versus instinct; it is time we learn not to speak of emotion versus cognition (see also MILLER/KELLER 2000).
Absence of neural perspectives
As I mentioned above, whether one considers the evidence for modularity to be obvious or underwhelming is a matter of opinion, reflecting the rigidity with which one holds to which definition. Again, P&P do not decry modularity in general (pun noted but not intended); rather, they find no evidence for modularity of cognitions (‘contra’ emotions) and of cortical pathways (contra subcortical pathways).
Certainly there is less evidence for modularity at the cortical level than at the subcortical (CAMHI 1984; PANKSEPP 1998). But this too, is neither a new finding nor a new debate (POLSTER/NADEL/SCHACTER
Evolution and Cognition ❘ 16 ❘ 2001, Vol. 7, No. 1
1991), and ‘less evidence’ of cortical modularity doesn’t mean ‘no evidence’: there is room here for empirical investigation (see ARBIB/ERDI 2000 and PAGE 2000). Cortical localization of function has been documented from many studies of human brain damage (e.g., COSMIDES/TOOBY 2000; GRODZIN-SKY 2000; HART/BERNDT/CARAMAZZA 1985; KOLB/ WISHAW 1990; SACKS 1985; TRANEL/DAMASIO 1985), as well as in intact, non-human animals (e.g., KEN-DRICK/BALDWIN 1987). Whether these specialized circuits provide inputs or outputs or both, or are considered to be processing and producing affects or cognitions, are still questions that are open to debate. Is face recognition purely cognitive? Is it an input or is it an output? What about recognition of facial expressions?
Anti-organic bias
These questions inevitably lead P&P to reframe both emotions and cognitions in terms of organic, rather than computational representations. I think this is important to do. While there are likely to be truly computational processes in the brain/mind (e.g., GALLISTEL 1995), there are also likely to be analog processes (e.g., CARMAN/WELCH 1992) and truly organic ‘embodied’ representations (see HUMPHREY 1992). I suspect that the computational metaphor, though widely used by evolutionary psychologists, is likely to be just metaphor much of the time.
But even if the metaphor is literally true more often than not, computation is not sufficient. I suspect that P&P are correct when they assert that “the foun-
dations of mind are fundamentally ‘embodied’ by organic processes that are impossible to compute except in the most superficial way”. Once again, however, this is an old debate which won’t be settled any time soon (see e.g., FODOR 1981; SEARLE 1981, 1990; WELLS 1998).
What Penance?
The terms ‘environment of evolutionary adaptedness’, ‘encapsulated modules’, ‘computations’ and for that matter, ‘representations’—were put forth as short-hand notations for things that we do not yet fully understand and therefore cannot yet adequately describe. Like all metaphors and analogies, they have their limits—and while we do need to take care not to push those limits and still believe that we are speaking ‘truth’, I think that their worth must be assessed not on their ultimate ability to ‘reflect’ reality, but on their practical value. If their use encourages inquiry, they are a good thing; if they discourage inquiry, they are a bad thing.
On the whole, I think such metaphors have encouraged inquiry and that they are, therefore, useful (see e.g., MEALEY/DAOOD/KRAGE 1996; MURPHY/ STICH 2000). DARWIN’s entire book “On the Origin of Species by Natural Selection” (1859) was an analogy, and although that analogy led to several misunderstandings—including Herbert SPENCER’s ‘survival of the fittest’—we nevertheless, still find value in DARWIN’s choice of words. The concept of a gene as a ‘particulate entity’ was, too, an analogy—an analogy that may have led MENDEL to overlook the reality of linkage—but an analogy which we still use today.
Debates about terminology permeate all disciplines, including other subdisciplines of evolutionary biology (e.g., GHISELIN 1981) and psychology (e.g., WIDIGER/SANKIS 2000). As elsewhere, the metaphors of modern evolutionary psychology will
References
- Alcock, J. (1998) Unpunctuated equilibrium in the Natural History essays of Stephen J. Gould. Evolution and Human Behavior 19: 321–336.
- Arbib, M. A./Erdi, P. (2000) Precis of ‘Neural Organization: Structure, Function, and Dynamics’. Behavioral and Brain Sciences 23: 513–571.
- Buss, D. M. (1995) Evolutionary psychology: A new paradigm for psychological science. Psychological Inquiry 6: 1–30.
- Buss, D. M./Haselton, M. G./Shackelford, T. K./Bleske, A. L./ Wakefield, J. C. (1998) Adaptations, exaptations, and spandrels. American Psychologist 53: 533–548.
Camhi, J. M. (1984) Neuroethology. Sinauer: Sunderland MA.
- Carman, G. J./Welch, L. (1992) Three dimensional illusory contours and surfaces. Nature 360: 585–587.
- Cosmides, L./Tooby, J. (1997) The modular nature of human intelligence. In: Scheibel, A. B./Schopf, J. W. (eds) The Origin and Evolution of Intelligence. Jones & Bartlett: Boston MA, pp. 71–101.
- Cosmides, L./Tooby, J. (2000) Social exchange: Converging evidence for special design. Presented at the Human Behavior and Evolution Society, Amherst College, Amherst MA, June 2000.
- Daly, M./Wilson, M. (1999) Human evolutionary psychology and animal behaviour. Animal Behaviour 57: 509–519.
- Damasio, A. R. (1994) Descarte’s Error: Emotion, Reason and the Human Brain. Putnam: New York.
- Damasio, A. R. (1999) The Feeling of What Happens: Body and Emotion in the Making of Consciousness. Harcourt Brace: New York.
- Darwin, C. (1859) On the Origin of Species by Natural Selection. John Murray: London.
- Eimas, P. D. (1985) The perception of speech in early infancy. Scientific American 252: 46–52.
- Eimas, P. D./Miller, J. L. (1992) Organization in the perception of speech by young infants. Psychological Science 3: 340–345.
- Ekman, P./Davidson, R. J. (eds) (1994) The Nature of Emotion: Fundamental Questions. Oxford University: New York.
- Finlay, B. L./Darlington, R. B. (1995) Linked regularities in the development and evolution of mammalian brains. Science 268: 1578–1584.
- Fodor, J. A. (1981) The mind–body problem. Scientific American 244: 114–123.
- Fodor, J. A. (1983) The Modularity of the Mind. MIT Press: Cambridge MA.
- Fodor, J. A. (2000) Why we are so good at catching cheaters. Cognition 75: 29–32.
- Gallistel, C. R. (1995) The replacement of general-purpose theories with adaptive specializations. In: Gazzaniga, M. S. (ed) The Cognitive Neurosciences. Bradford: Cambridge MA, pp. 1255–1267.
- Gazzaniga, M. S. (1989) Organization of the human brain. Science 245: 947–952.
- Ghiselin, M. T. (1981) Categories, life, and thinking. Behavioral and Brain Sciences 4: 269–313.
- Goode, E. (2000) Human nature: Born or made? Evolutionary theorists provoke an uproar. NY Times, March 14, 2000, pD1, D9.
- Gould, S. J. (1991) Exaptation: A crucial tool for evolutionary psychology. Journal of Social Issues 47: 43–65.
- Gould, S. J./Lewontin, R. C. (1979) The spandrels of San Marco and the Panglossian paradigm: A critique of the adapta-
tionist programme. Proceedings of the Royal Academy of London, B 205: 581–598.
- Griffiths, P. E. (1990) Modularity, and the psychoevolutionary theory of emotion. Biology and Philosophy 5: 175– 196.
- Griffiths, P. E. (1997) What Emotions Really Are: The Problem of Psychological Categories. University of Chicago: Chicago IL.
- Grodzinsky, Y. (2000) The neurology of syntax: Language use without Broca’s area. Behavioral and Brain Sciences 23: 1– 71.
- Hall, B. K (ed) (1994) Homology: The Hierarchical Basis of Comparative Biology. Academic: San Diego CA.
- Hart, J./Berndt, R. S./Caramazza, A. (1985) Category-specific naming deficit following cerebral infarction. Nature 316: 439–440.
- Harvey, P. H./Purvis, A. (1991) Comparative methods for explaining adaptations. Nature 351: 619–624.
- Hauser, M. D. (1996) The Evolution of Communication. Bradford: Cambridge MA.
- Hauser, M. D. (2000) Why humans are the wrong species in which to study the evolution of human intelligence. Plenary Address to the Human Behavior and Evolution Society, Amherst College, Amherst MA, June 2000.
- Hinde, R. A. (1966) Animal Behaviour: A Synthesis of Ethology and Comparative Psychology. McGraw-Hill: New York.
- Humphrey, N. (1992) A History of the Mind: Evolution and the Birth of Consciousness. Simon & Schuster: New York.
- Jacob, F. (1977) Evolution and tinkering. Science 196: 1161– 1166.
- Jaynes, J. (1969) The historical origins of ‘Ethology’ and ‘Comparative Psychology’. Animal Behaviour 17: 601– 606.
- Hirsch, J. (ed) (1987) Comparative Psychology: Past, Present, and Future. Journal of Comparative Psychology 101: 219– 292.
- Kendrick, K. M./Baldwin, B. A. (1987) Cells in temporal cortex of conscious sheep can respond preferentially to the sight of faces. Science 236: 448–450.
- Kolb, B./Wishaw, I. Q. (1990) Fundamentals of Human Neuropsychology. 3rd edition. W. H. Freeman & Co.: San Fansisco CA.
- Larson, A./Losos, J. B. (1996) Phylogenetic systematics of adaptation. In: Rose, M. R./Lauder, G. V. (eds) Adaptation. Academic: New York, pp. 187–220.
- Lazarus, R. S. (1991) Emotion and Adaptation. Oxford University: Oxford.
- Lenneberg, E. (1967) The Biological Foundations of Language. John Wiley & Sons: New York.
- Liberman, A. M./Mattingly, I. G. (1989) A specialization for speech perception. Science 243: 489–494.
- Lieberman, P. (1977) The phylogeny of language. In: Sebeok, T. A. (ed) How Animals Communicate. University of Indiana: Bloomington IN, pp. 3–25.
- MacLean, P. D. (1990) The Triune Brain in Evolution: Role in Paleocerebral Function. Plenum: New York.
- Mallon, R./Stich, S. P. (2000) The odd couple: The compatibility of social construction and evolutionary psychology. Philosophy of Science 67: 133–154.
- Mealey, L. (1994) Review of ‘The Adapted Mind: Evolutionary Psychology and the Generation of Culture’, edited by J. Barkow, L. Cosmides, and J. Tooby. Politics and the Life Sciences 13: 294–295.
- Mealey, L. (1996) Evolutionary psychology: The search for evolved mechanisms underlying complex human behavior. In: Hurd, J. (ed) The Biology of Morality. Edwin
Evolution and Cognition ❘ 18 ❘ 2001, Vol. 7, No. 1
Mellen: Lewiston NY, pp. 35–61.
- Mealey, L. (2000) Sex Differences: Developmental and Evolutionary Strategies. Academic Press: San Diego CA.
- Mealey, L. (2001) What? Me worry? The state of human ethology in the year 2000: Presidential Address to the International Society for Human Ethology. The Human Ethology Bulletin 15: 2–8.
- Mealey, L. (in press) Kinship: The ties that bind (disciplines). In: Holcomb III, H. R. (ed) EConceptual Challenges in Evolutionary Psychology. Kluwer Academic Publishers: New York.
- Mealey, L./Daood, C./Krage, M. (1996) Enhanced memory for faces associated with potential threat. Ethology and Sociobiology 17: 119–128.
- Miller, G. A./Keller, J. (2000) Psychology and neuroscience: Making peace. Current Directions in Psychological Science 9: 212–215.
- Miller, G. F. (2000a) Alas, poor scholarship. The Evening Standard (London), July 10. Retrieved from the World Wide Web: http://www.thisislondon.co.uk/dynamic/lifestyle/review.html?in_review_id=295797& in_review_text_id=240358
- Miller, G. (2000b) How to keep our metatheories adaptive:
- Beyond Cosmides, Tooby and Lakatos. Psychological Inquiry 11: 42–46.
- Murphy, D./Stich, S. (2000) Darwin in the madhouse: Evolutionary psychology and the classification of mental disorders. In: Carruthers, P./Chamberlain, A. (eds) Evolution and the Human Mind: Modularity, Language and Meta-Cognition. Cambridge University: Cambridge, pp. 62–92.
- Nesse, R. (1991) What is mood for? Psycoloquy (an electronic journal) 2.9.2.1.
- Nesse, R. (1992a) Overevaluation of the mood-emotion distinction. Psycoloquy (an electronic journal) 3.2.1.2.
- Nesse, R. (1992b) Ethology to the rescue (Reply to Plutchik). Psycoloquy (an electronic journal) 3.2.1.6.
- Page, M. (2000) Connectionist modelling in psychology: A localist manifesto. Behavioral and Brain Sciences 23: 443– 512.
- Panksepp, J. (1982) Toward a general psychobiological theory of emotions. Behavioral and Brain Sciences 5: 407–467.
- Panksepp, J. (1998) Affective Neuroscience. Oxford University Press: New York.
- Pinker, S. (1994) The Language Instinct. Wm Morrow: New York.
- Pinker, S. (1997a) Language as a psychological adaptation. In: Bock, G. R./Cardew, G. (eds) Characterizing Psychological Adaptations. Wiley: Chichester, pp. 162–172.
- Pinker, S. (1997b) How the Mind Works. Norton: New York. Pinker, S./Bloom, P. (1990) Natural language and natural se-
- lection. Behavioral and Brain Sciences 13: 707–784. Plutchik, R. (1980) Emotion: A Psychoevolutionary Synthesis.
- Harper & Row Pub: New York. Plutchik, R. (1992) What is mood for? A critique. Psycoloquy
- (an electronic journal) 3.2.1.5. Polster, M. R./Nadel, L./Schacter, D. L. (1991) Cognitive neuroscience analyses of memory: A historical perspective. Journal of Cognitive Neuroscience 3: 95–116.
- Ramachandran, V. S./Blakeslee, S. (1998) Phantoms in the Brain. Wm Morrow: New York.
- Rolls, E. T. (2000) Precis of ‘The Brain and Emotion’. Behav-
ioral and Brain Sciences 23: 177–234.
- Sacks, O. (1985) ‘The Man Who Mistook his Wife for a Hat’ and Other Clinical Tales. Summit: New York.
- Schleidt, W. M. (1974) How “fixed” is the fixed action pattern? Zeitschrift fur Tierpsychologie 36: 184–211.
- Searle, J. R. (1981) Minds, brains and programs. In: Haugeland, J. (ed) Mind Design: Philosophy, Psychology, Artificial Intelligence. Bradford: Cambridge MA, pp. 282–306. Reprinted from Searle (1980) Behavioral and Brain Sciences 3: 417–424.
- Searle, J. R. (1990) Consciousness, explanatory inversion, and cognitive science. Behavioral and Brain Sciences 13: 585–642.
- Seligman, M. E. P. (1970) On the generality of the laws of learning. Psychological Review 77: 407–418.
- Shallice, T. (1988) From Neuropsychology to Mental Structure. Cambridge University Press: Cambridge.
- Shallice, T. (1991) Precis of ‘From Neuropsychology to Mental Structure’. Behavioral and Brain Sciences 14: 429–469.
- Skinner, B. F. (1981) Selection by consequences. Science 213: 501–504.
- Thiessen, D. T. (1998) Expanding the boundaries of evolutionary psychology: The context of domain-specific adaptations. Mankind Quarterly 38: 337–362.
- Thornhill, R. (1997) The concept of an evolved adaptation. In: Bock, G. R./Cardew, G. (eds) Characterizing Psychological Adaptations. Wiley: Chichester, pp. 4–13.
- Tooby, J. (1999) The most testable concept in biology, Part 1. Human Behavior & Evolution Society Newsletter 8: 1–4.
- Tooby, J./Cosmides, L. (1990) The past explains the present: Adaptations and the structure of ancestral environments. Ethology and Sociobiology 11: 375–424.
- Tooby, J./Cosmides, L. (1992) The psychological foundations of culture. In: Barkow, J./Cosmides, L./Tooby, J. (eds) The Adapted Mind. Oxford University: New York, pp. 19–136.
- Tranel, D./Damasio, A. R. (1985) Knowledge without awareness: An automatic index of facial recognition by prosopagnosics. Science 228: 1453–1454.
- Van Parijs, P. (1981) Evolutionary Explanation in the Social Sciences. Rowman & Littlefield: Totowa NJ.
- Wells, A. (1998) Evolutionary psychology and theories of cognitive architecture. In: Crawford, C./Krebs, D. L. (eds) Handbook of Evolutionary Psychology: Ideas, Issues and Applications. Lawrence Erlbaum: Mahwah NJ, pp. 235– 264.
- Werker, J. F./LaLonde, C. E. (1988) Cross-language speech perception: Initial capabilities and developmental change. Developmental Psychology 24: 672–683.
- Widiger, T. A./Sankis, L. M. (2000) Adult psychopathology: Issues and controversies. Annual Review of Psychology 51: 377–404.
- Wilson, D. S. (1994) Adaptive genetic variation and human evolutionary psychology. Ethology and Sociobiology 15: 219–235.
- Young, J./Persell, R. (2000) On the evolution of misunderstandings about evolutionary psychology. Annals of the New york Academy of Science 907: 218–223.
- Zatorre, R. J./Evans, A. C./Meyer, E./Gjedde, A. (1992) Lateralization of phonetic and pitch discrimination in speech processing. Science 256: 846–849.
Affective Neuroscience, Psychiatry, and Sociophysiology
Affective Neuroscience
My reaction to PANKSEPP and PANKSEPP (2000) reflects the viewpoint of a U.S. psychiatric educator, clinician and researcher who finds their approach highly compatible with a needed nascent basic science framework of the psychiatric medical specialty, as well as its allied human clinical services disciplines. This sociophysiological framework for understanding the pathogenesis of psychiatric illness (GARDNER 1996, 1997) depends in part on the concept of basic plans for communicational behavior with conspecifics (members of the same species) originating in ancient brain structures hundreds of millions of
Abstract
The PANKSEPPs laudably emphasize that evolutionary psychology (EP) has not appreciated the ancient contributions to the human brain; rather EP conflates subcortical emotional factors with cognitions amplified in recent human evolution. Remarkably, despite overvaluing the mushroomed cortex, EP pays little attention to communicative and social facets of human development although new work shows that a significant proportion of the variance in primate brain size stems from its correlation with species-group size. Jaak PANKSEPP’s earlier work on emotional communications assists the development of a basic science of psychiatry, a framework now sadly lacking. Instead, market forces in the absence of a comprehensive scientific rationale have led to what could be summarized as ‘relationshipless’ values that are contrary to the ally-formation particularly facilitated by human brains. In concert with PANKSEPPs’ argument, psychiatry requires a basic science framework titled sociophysiology entailing comparisons (similarities) as well as contrasts of humans to other animals.
Key words
Emotion, communication, sociophysiology, psychiatry.
years ago. The PANKSEPPs document that evolutionary psychologists (EP) contrastingly focus on recently evolved modular cognition as the central brain advance, a focus encouraged in part by the impressive cortical specializations seen by SPERRY and GAZZANIGA in people with operatively caused ‘split-brains’. No-one denies that the human cerebral cortex is a most impressive organ, but the PANK-SEPPs point out that important human experiences—emotions—reflect the dominating actions of subcortical structures, including those originating in deep time, not just recently expanded adaptations less than a few hundred-thousand years old.
Emotional expressions reflect communicational basic plans so that the PANKSEPP focus complements the sociophysiological framework.
They highlight the problems by suggesting problems in EP thinking are “sins”. Making definitive comments but using mildly humorous imagery holds no surprise because Jaak PANKSEPP’s most recent striking research findings show play and laughter to be much more primitive mammalian communications than heretofore suspected. With BURGDORF, he used the infant tickle response in the rat pup as the means of exploring this communicative interaction well known to humans (PANKSEPP/BURG-DORF 2000). They col-
lected data and drew conclusions with great care, paying attention to all the alternative hypotheses that they, colleagues and critics could generate. This cautious methodology lends additional power to the other arguments Jaak PANKSEPP makes.
Elsewhere he emphasized the subcortical location of neural systems that mediate seven emotion-systems; he investigated, consolidated and documented these in his extraordinary book, Affective Neuroscience (PANKSEPP 1998). Armed with original research done mostly with rat subjects, he has persuasively argued for the existence of emotions in non-human animals and also for the relevance of
Evolution and Cognition ❘ 25 ❘ 2001, Vol. 7, No. 1
these subcortical systems to human consciousness and other facets of thought (PANKSEPP 2000). I feel the PANKSEPP approach fosters integration between the behavioral–mental–experiential (BME) and the organ–cellular–molecular (OCM) levels of analysis. His work exemplifies scientific “docking” between the levels.
By contrast, the EP thinkers fundamentally restrict their thinking to the BME level. They use the word biology to suggest otherwise, but many of these thinkers refer little if at all to actual brain research. Rather, they focus primarily on parameters of behavior that they imagine solidified in a Pleistocene Era of Evolutionary Adaptedness resulting from genome changes that led to humans becoming distinct from other animals, with other functions held in common such as eating and defense taken for granted and less interesting. Philip LIEBERMAN (2000), who has benefited from Paul MACLEAN’s formulations (MACLEAN 1990), argued in another extraordinary recent book that even the human specialty, verbal language, stems in large part from subcortical mechanisms, rather than reflecting a bureaucracy implicitly limited to the cerebral cortex as popularized, by EP linguist Steven PINKER (1994), for instance. LIEBERMAN notes EP modular theorists use core metaphors from World War II electronics that infer direct sequential connections from module to module. But feed-forward and feedback connections found in all cortical areas directly counter the presupposed sequential connections.
Not only LIEBERMAN, but PANKSEPP too benefited from MACLEAN’s emphasis on evolutionary origins of present day brains that focused on complex behaviors involving other conspecifics. PANKSEPP worked directly with John Paul SCOTT who died in 2000 and to whom the PANKSEPPs dedicate their seven sins–paper. SCOTT pioneered the genetics of behavior, focusing in landmark studies on behavioral differences in dog breeds (SCOTT/FULLER 1965). Significantly SCOTT (1989) wrote on the evolution of human social systems.
Conceptually linking the BME and the OCE levels of analysis has commanded great interest but also incurred great problems over the two and a half millennia since the school of HIPPOCRATES defined the brain as the body’s control center. Typically, as with the EP workers focusing only on the BME level, either one or the other level has been featured in isolation, with the other revealed as trivial with only minimum information needed. For a recent example of a contrasting focus restricted to the OCM level, KANDEL and SQUIRE (2000) published an article with
an engaging title that included, “breaking down scientific barriers to the study of the brain and mind”. But in fact they considered only cognition and memory on the upper level and suggested that new advances in the dramatic breakthroughs in OCM research will illumine the BME level. Their tone showed little appreciation for the need of a two-way street —that conceptualizations at the BME level are required for docking between the two levels. Yet such are required for full meaning to such disciplines as psychiatry and related clinical human care-giving disciplines. Emergent properties at the more complex level—such as story-production—determines much behavior not predictable from features of the less complex level. Beyond its cognitive and memory aspects alone, conspecific communication represents a brain-driven activity that requires analysis. Contrasting to the KANDEL-SQUIRE approach, linguist PIATTELLI-PALMARINI (2000) opined recently in Nature that “The brain scientists will have to know exactly what they are expected to find the neural bases of”.
The PANKSEPPs’ “emotional” approach (pun intended) curiously returns us to what the HIPPOCRATIC School suggested when as the brain emerged as the major controlling center of the body during the 5th century B.C.: “Men ought to know that from nothing else but the brain comes the joys, delights, laughter and sports, and sorrows, griefs, despondency, and lamentations. And by this, in an especial manner, we acquire wisdom and knowledge, and see and hear and know what are foul and what are fair, what are bad and what are good, what are sweet and what are unsavory” (FINGER 2000). Note in this HIPPOCRATIC thinking, wisdom and knowledge take second place to affectively charged communications and feelings. I intended the above pun because I suspect that one of the barriers to considering fully the integration between the BME and OCM levels stems from strong feelings on the part of senior neuroscientists that emotions and other communicational topics interfere with proper science, extending to the idea that those who pay attention to them are necessarily unreliable, involved with fluff somehow, or willing to tolerate dubious information. MACLEAN, SCOTT and Jaak PANKSEPP represent a handful of pioneers who juxtapose emotions and other conspecific communications as meaningful topics that integrate the two levels. But KANDEL and SQUIRE in their listing of 20th century notables omit neuroscientists MACLEAN and PANKSEPP, though both made highly significant contributions, at least if BME–OCM integrative issues were valued.
Evolution and Cognition ❘ 26 ❘ 2001, Vol. 7, No. 1
Most psychiatrists have lived either on the BME level or the OCM. But the two levels should be juxtaposed, at least as an ideal. However, selective allegiances at one or the other conceptual level tell the tumultuous 20th century story of the medical specialty, which I will summarize briefly before moving on to how a group of psychiatrists and other scholars that I here represent have worked to remedy the situation. To preview these developments, the Across-Species Comparisons and Psychopathology (ASCAP) Society brainstormed for a decade, attempting appropriate conceptual docking between the BME and OCM levels. How can we rationally formulate psychiatric conditions and social distresses using the other medical specialties and their functional relations to their basic sciences as models? That is, a pathological condition is best diagnosed and treated when understood as a variant of the normal condition like a stroke from a damaged blood vessel can be diagnosed and treated best with knowledge of normal brain circulation. Sociophysiology represents the brain physiology of social and communicative processes—especially among conspecifics. The organ of interest surely cannot be limited to the cerebral cortex but involves all parts concerned with detection, analysis and actions of other people, though the cortex of course can hardly be ignored. The sociophysiological approach to such matters both compares (examines similarities between) humans and other animals and also contrasts them. The large cortex, we presume, results from its storyusing capacity (DONALD 1991), which provides great resourcefulness for therapy, whether this primarily represents pharmacotherapy, psychotherapy, both, or group and family variants.
20th Century Psychiatry
Freud, a sophisticated 19th century neurologist when he founded psychoanalysis, felt too little was known of the nervous system so he focused only on BME with the expectation that his work would eventually link fruitfully with the OCM (SULLOWAY 1992). He relied in his mind-models on machineimagery of the 19th century (RABINBACH 1990). His most important therapeutic tactic, however, involved transference, the idea that the patient behaves with the doctor in ways that reflect previous relationships. After the World War II exodus from Europe of distinguished psychoanalysts, this arena of thought and clinical activity became the U.S. mid-century academic standard although, not being evidentially based in the body, the approach did not exhibit parallelism with other medical specialties.
But in recent decades, the pendulum swung to the other extreme. A major shift in psychiatry stemmed in part from the anti-analytic movement led by biochemist–psychiatrist Eli ROBINS after he experienced damage from his too-authoritarian psychoanalyst. ROBINS and his articulate prolific followers fostered an increasingly data-based form of clinical activity with curious de-emphasis of pathogenesis. Psychoanalysis had featured pathogenesis with metaphors independent of brain-data; perhaps in response, the ROBINS-led psychiatrists eschewed pathogenesis as premature and to be done eventually by biochemists (an OCM level of analysis). They propounded that accurate description of disorders and their correlates and treatments should come first, including a fresh emphasis on the major disorders such as bipolar and schizophrenic conditions. They demanded that any clinical treatment should possess research validation. The movement omitted relevancy of normal behavior, focusing only on the pathological.
However, the American Psychiatric Association’s Diagnostic and Statistical Manual newly emerged in 1980 with codified diagnoses featuring operational definitions. This potentially helped BME–OCM integration because it fostered naturalistic descriptions. Moreover, psychopharmacology has helped many patients with serious maladies, and, as well, provides formulations of how the brain works with reference to anxious and affective states as well as clarifying behaviorally and thought-disordered states. The OCM level of analysis greatly benefited patients. But psychiatry struggles.
From outside the field, distinguished physiologist–anthropologist, Jared DIAMOND, commented recently on academic psychiatry in a “think tank” column of the New York Times (DIAMOND 2001). Echoing my own impressions and those of many psychiatrist colleagues, DIAMOND states, “[A] toonarrow focus on biological psychiatry … hurts patients. I fully accept the importance of biological psychiatry, having devoted some of my own research to problems in that area … But now the problem has swung to the opposite extreme: psychiatry departments have become bastions of molecular psychiatry, at which more time is devoted to studying and teaching psychopharmacology than to what are called talk therapies”. DIAMOND goes on to conclude about transference that “Understanding [the patient–doctor] unique relationship was one of the… insights that put Freud right up there with DARWIN”.
Evolution and Cognition ❘ 27 ❘ 2001, Vol. 7, No. 1
The enormous swing in psychiatric practice augmented by major changes in payment schemes produced powerful distressing repercussions, documented by anthropologist Tanya LUHRMAN in her book, “Of Two Minds: The Growing Disorder in American Psychiatry” (2000). In the course of think tank deliberations in the Research Committee at the Group for the Advancement of Psychiatry (GAP), academic child psychiatrist Karen Dineen WAGNER suggested that the ideal for psychopharmacology and managed care has become “relationshipless psychiatry”. Examiners for Board Certification at the American Board of Psychiatry and Neurology feel that interpersonal skills are seriously lacking in many observed candidates. In conclusion, there now exist increasingly high-level awarenesses both in and out of the field that psychiatry needs a core model for how the brain at its center operates. The above-mentioned Research Committee of the GAP suggested that such a model would be “the social brain” (BAKKER et al. 2001).
Sociophysiology
I turn now to my compatible but different intellectual route from that of Jaak PANKSEPP. My background as an academic psychiatrist included research on the ethology of sleep movement (GARD-NER/GROSSMAN 1975) and evolved to a role as educator/administrator. In the course of this, an assignment as chairman of a medical school’s curriculum committee affected my thinking. The responsibility occurred during a major curricular revolution and involved a charge from the dean to increase hours for educating medical students in “behavioral science,” a national trend. I of course supported these changes because behavioral science issues and interviewing skills provided foundations for all future physicians, not psychiatrists only. But even as I fostered the major changes successfully, I also felt cognitive dissonance because I had been particularly interested in medical pathogenesis since medical school and saw no evidence my fellow educators showed understanding of this for psychiatry, nor, for that matter, did my psychiatrist colleagues, even those interested in education. No tradition existed for understanding psychiatric symptoms or illnesses as deviations from normal brain processes, as symptoms of congestive heart failure represent the heart muscle’s more limited ability. I realized that psychiatry’s basic science needed articulation and started with the alpha communications typical of a manic patient (GARDNER 1982). PRICE (1967) well before me had suggested that ancient biology of social rank hierarchy plays significant roles in affective illness.
I found with Joan and Carl GUSTAVSON that operationally defined “manic” communications did not differ from those of charismatic leaders (GARDNER et al. 1985). This resonated with the idea that an alpha basic plan likely underlay the behavior, and given similarities of these communications to those of dominant animals, represented factors transmitted via ancient genes determining brain structure. With the concept of propensity states antedating language in communication (psalic), I proposed that communication and sociality foster important ancient brain states retained in the present (GARDNER 1988, 1998a). Signaling the “planful” attributes of living matter emphasized by MAYR (1982), psalic also refers to programmed spacings and linkages in conspecifics, fundamental aims of communication. Particular psalics take definition from existing in (1) normal humans, (2) psychiatrically disturbed humans, and (3) animals. Two psalics include alpha psalic, of course, seen in mania, normal leadership, and animal dominance, and the related audience psalic (state of receptivity to conspecifics as in cult membership, normal audiences, and animal subordination). Sample others include mating, nurturant and nurturance-eliciting psalics.
Molecular biologist C. U. M. SMITH (1993) wrote an editorial supportive of the psalic concept for the British Journal of Psychiatry. Support for basic plan approaches stem now from recent neurobiological research. KANDEL and SQUIRE’s neuroscience review (2000) cited the following data-based advances: “molecular machinery and electrical signaling properties of neurons are widely conserved across animal species,” that “what distinguishes one species from another, with respect to their cognitive abilities, is the number of neurons and the details of their connectivity,” and that “remarkable principles of evolutionary conservatism are emerging from the study of nerve cells”.
In 1987, I commenced publication of The Across-Species Comparisons and Psychopathology (ASCAP) Newsletter that represented a brainstorming operation. It ran monthly for twelve years with an international circulation and now continues on a quarterly basis as The ASCAP Bulletin. The ASCAP publication also now additionally serves as the communicational organ for the Psychotherapy Section of the World Psychiatric Association. The ASCAP Society commenced in 1991, its first president ethologist Michael CHANCE, and meets at least yearly. AS-
Evolution and Cognition ❘ 28 ❘ 2001, Vol. 7, No. 1
CAP activities include conferring the Aaron T. Beck Award to the most deserving essay on evolutionary biology and psychopathology. Membership disciplines and roles include psychiatry, ethology, psychology, sociology, social work, literature, philosophy, anthropology, animal behavior, veterinary science, sociobiology, psychotherapy, and psychiatric education, specifically, residency directorship and department chairmanship. Formerly, a number of evolutionary psychologists also counted amongst the ranks (indeed some served as president), but some have dropped away, sensing, I believe, that basic plans conceptions remain valued by the core members of the society. Thus, from its beginnings as the organ of the ASCAP Society, the mission statement has read as follows:
“The ASCAP Society represents a group of people who view forms of psychopathology in the context of evolutionary biology and who wish to mobilize members and resources of various disciplines so as to enhance the further investigation and study of the conceptual and research questions involved. This scientific society is concerned with the basic plans of behavior that have evolved over millions of years and that have resulted in psychopathologically related states. We are interested in the integration of various methods of study ranging from cellular processes to individuals in groups.”
Publications in refereed journals that do not reflect an EP slant have resulted from the newsletter’s brain-storming efforts. Examples include a report on PRICE’s hierarchical theory of affective illnesses published initially in the British Journal of Psychiatry (PRICE et al. 1994) and reprinted in an edited book. Another often cited report was published in the American Journal of Psychotherapy (SLOMAN et al. 1994). Textbooks on evolutionary or DARWINIAN psychiatry have now emerged, written by ASCAP members (STEVENS/PRICE 1996; MCGUIRE/TROISI 1998). A book entitled Genes on the Couch (GILBERT/BAILEY 2000) stemmed from the pages of The ASCAP Newsmatters by making the following points: people both contrast and compare to other animals. We compare (are similar) to them in that we possess many of the same needs and instincts, as say, one’s dog. Other animals have emotions (pet owners know PANKSEPP to be correct). Moreover, we shiver with cold as do other animals and experience other automatic reactions that stem from lower brain levels. However, we also contrast in having a larger brain, one indeed three times heavier than the chimpanzee brain despite nearly identical genomes—1.6% difference is commonly cited—and the human cerebral cortex flattened would cover an area four times greater than a chimp cortex would (CALVIN 1990). What is that big brain for? Following DUNBAR (1996), we concluded that allies, thinking and planning (ATP) are key helpful attributes, with allies chief among these (contrary to EP thinking). If one has shiver-like responses that cause trouble (impulsivity, rage, anxiety), one should look for allies, who then aid thinking and planning. Indeed we see allies as not only physicians and therapists but also 12-step programs, anger management groups, school counselors and innumerable other cultural or societal institutions. Humans rely on strangers as well as kin. I have proposed that we possess a peoplefocused “story-using brain” (probably dependent on cortex) that also facilitates such conspecific benefits (GARDNER 1998b).
I believe that these represent understandable, unfanciful, common-sensical and data-based explanations. Patients find them helpful. They do not supplant other psychotherapies, but rather augment them. Other psychotherapies, may, however, benefit from sociophysiological analysis (GARDNER 2001).
In summary, I agree that EP has underemphasized the ancient contributions to the human brain and conflated emotional factors with cognitions augmented by recent human evolution. We welcome Jaak PANKSEPP’s work on emotional communications as complementary to sociophysiology’s develop-
letter and from the 1997 AS-CAP Society annual conference.
To illustrate how sociophysiology works in clinical practice, I regularly explain to patients about evolutionary
Degrees of Modularity
HE TARGET ARTICLE BY Panksepp and Panksepp (P&P) is about the holy grail of the psychological sciences: to generate a description of the human mind that can be mapped on the description of the neural circuitry studied by neuroscientists. P&P’s question is whether evolutionary psychology, in the form that emerged during the 1990s, is a good starting point for this grand synthesis and whether it is at all compatible with what we already know about the brain. T
The variety of evolutionary psychology criticized by P&P is based in large part on the writings of Leda COSMIDES and John TOOBY. Its core assumption is the principle of massive modularity, also known as the Swiss Army Knife model. It postulates dozens to hundreds of special-purpose
Abstract
It is argued that the distinction between cognitive and modular functions as originally proposed by J. FODOR (1983) is valid, but that two types of modular functions should be distinguished: (1) hard-wired ‘primary’ modules that are mainly but not exclusively subcortical and are mainly concerned with motivation (e.g. fear conditioning); and (2) ‘modularized’ functions that are mainly cortical, are shaped by recurrent sensory and/or cognitive input, and are concerned mainly with sensory analysis and motor control (e.g. language). A strict distinction between hard-wired subcortical circuits and non-modular cortical functions, as proposed by Panksepp and Panksepp, is rejected. Also the ‘Swiss army knife model’ of evolutionary psychology is held to be simplistic. Instead it is proposed that many brain mechanisms, including the cognitive system, are multi-purpose. These ‘guns for hire’ are recruited by functionally specialized modules to produce adaptive behavior. It is argued that the most useful conceptual framework is an expanded ethological model in which specialized brain systems may use either sensory or cognitive input and may produce either behavioral or cognitive output.
Key words
Evolutionary psychology, modularity, cognitive system, exaptation.
mechanisms, each ‘designed’ by evolution to solve an adaptive problem (KENRICK/SADALLA/KEEFE 1998; PINKER 1994; TOOBY/COSMIDES 1992). According to FODOR (1983), a module is a functionally specialized processing device. It is informationally encapsulated, working only on a narrow set of inputs specified by its input connections; it produces a specialized, stereotyped, ‘shallow’ output; it is hard-wired, developing during ontogenesis without the need for effortful learning; and its inner workings are not accessible to introspection: it is cognitively impenetrable.
The special-purpose devices of COSMIDES and TOOBY have the essential properties of FODOR’s modules, with the additional specification that they evolved as adaptive responses to challenges that were repeatedly encountered in ancestral environments. However, for FODOR modularity was limited to the sensory and motor periphery of the mind while the cognitive system proper was nonmodular. COSMIDES and TOOBY have dropped this distinction by claiming that also cognition itself is domain-specific, specialpurpose, and effectively modular. The prominent examples for this view are CHOMSKY’s theory of universal grammar, recently popularized by Steven PINKER (PINKER 1994), and the finding that deontic reasoning routines—that is, reasoning about social interactions—differ from
reasoning in other domains in adaptively meaningful ways (COSMIDES/TOOBY 1992).
FODOR drew his examples of modularity from the sensory and motor systems which are in large part cortical. The modular systems that P&P are interested in are special-purpose subcortical circuits that are concerned with motivation and emotion. P&P insist that the closest we have to evolved specialpurpose devices for dealing with adaptive challenges are the subcortical mechanisms that generate action tendencies, emotions, and affective states. They offer what looks like a dichotomy between subcortical modular systems and the developmentally malleable ‘association cortex’.
This dichotomy is too simple to be useful. There are various types of functional organization in the vast expanses of association cortex. First, there is the cognitive system proper that FODOR opposes to the modules of the sensory–motor periphery. The core constituent of the cognitive system is working memory, with short-term memory buffers and executive control systems to manipulate the information that is held in the buffers (BADDELEY/LOGIE 1999). Presumably, the neuronal populations that form the buffers are able to respond with sustained activity changes to external stimuli (GOLDMAN-RAKIC 1995). The information held in the buffers at any one time can be described as a mental representation, a mental model, or a cognitive map, depending on the research tradition and the theorist’s preferences. This information can be entered into and retrieved from long-term declarative memory through the medial temporal lobe system. Presumably the mental representations in the buffers of working memory are the only elements of the mind that are consciously experienced and accessible to introspection (BADDELEY /ANDRADE 1998). Modules cannot be introspected because they keep no records of their activities. Intelligence is best defined as the ability to manipulate the information that is held in the buffers of working memory (KYLLONEN/CHRISTAL 1990).
Association cortex is also involved in rapid, nonconscious sensory–motor processing. The elaboration of visually-guided movements in areas of the parietal cortex is the most thoroughly studied example (BATISTA et al. 1999; COLBY/DUHAMEL 1996). These functions are not ‘primary’ modules of the kind that FODOR and the evolutionary psychologists have in mind because they are not strictly hardwired. They are learned by a neural substrate that is strategically located and structurally pre-designed for this kind of learning. We can call them modularized, as opposed to truly modular functions, or we can call them ‘secondary’ modules if we like. Except for their learned rather than hard-wired nature, secondary modules have most or all of the properties that FODOR uses to define (primary) modules.
Modularized functions may have been learned with conscious effort before becoming modularized. This is certainly the case for car-driving routines. Studies on eye movement conditioning in the parietal cortex of monkeys have also produced evidence for reinforcement systems that may possibly be used to guide learning in the absence of cognitive input (PLATT/GLIMCHER 1999). I agree with P&P that some of the modular functions that are commonly cited by evolutionary psychologists, including the classical example of language, are modularized rather than truly modular.
Even many of the instinctive behaviors studied by ethologists depend on learning. Thus, in classical imprinting the sensory template on which the key stimulus is fitted can mature only with proper sensory input. The goslings that became imprinted on Konrad LORENZ (LORENZ 1935) were born with a crude sensory template that specified no more than a large moving object that makes noises. What exactly mother goose looks like has to be learned. In this case the motor output is quite rigid, but the sensory template has to be fleshed out through learning.
In the case of language, a rather pliable neural substrate is strategically located between the auditory cortex and the motor outputs to the vocal apparatus. This neural substrate seems to be pre-formatted for making categorical distinctions between sounds, words, and word meanings, and for recognizing hierarchical relationships. In deaf-mutes this same substrate imposes categorical distinctions and hierarchical relationships on visual inputs during the learning of sign language (NEVILLE et al. 1998; NISHIMURA et al. 1999), in apparent violation of FODOR’s definition that modules are informationally encapsulated. That we are dealing with fairly general properties of the neural substrate, rather than the unique features of a language module, is also suggested by the observation that compared to righthemisphere thinking, left-hemisphere thinking in general depends on categorical distinctions, bounded entities, and hierarchical relationships between these entities (CORBALLIS 1991; DEGLIN/KINS-BOURNE 1996). This is the reason why the left hemispheres that write target articles and commentaries try so hard to define categorical distinctions between subcortical circuits and association cortex, or between modular and modularized systems although in reality we are most likely dealing with a continuum. There may be few if any brain functions that are 100% hard-wired or 100% arbitrarily learned.
It is possible that there is a tendency for undifferentiated cortex to evolve into hard-wired modules. The primate visual system has a complex hard-wiring with at least 30 ‘centers’ that are involved in specialized functions such as motion detection, color vision, shape recognition, face recognition, and the recognition of biological movements (VAN ESSEN/ DEYOE 1995). We may speculate that this vast system, which is now largely hard-wired, evolved from a poorly-differentiated substrate that originally per-
Evolution and Cognition ❘ 32 ❘ 2001, Vol. 7, No. 1
formed visual analysis in a modularized, rather than modular fashion.
P&P claim that not a single sociobiological module has ever been demonstrated at the cortical level. However, there are face-selective cells in the ventral visual stream of the temporal neocortex and related brain areas that develop within months after birth (RODMAN/SCALAIDHE/GROSS 1993), probably from a crudely pre-formed circuitry that is fine-tuned by imprinting-like learning. Even newborns prefer looking at faces as compared to other equally complex stimuli (GOREN/SARTY/WU 1975; MAURER 1985). The face cell circuitry and its behavioral manifestations appear too early in development to be produced by ‘nothing but’ recurrent sensory input and reinforcement history. There are also systems for smiling and other emotional expressions in the cingulate cortex (DAMASIO 1994, pp139–143; FRIED et al. 1998). Babies start smiling at faces by about 3 months of age. It is reasonable to hypothesize that this smile reflex involves the face cells of the ventral temporal lobe, the smile circuitry in the cingulate cortex, and hypothetical projections from the face cells to the ‘smile center’. The three-month delay after birth for the onset of smiling is not necessarily due to the need for learning by an undifferentiated neural substrate. It may reflect the gradual maturation of one or another component in an essentially hard-wired system. Hard-wired brain circuitry need not be present at the time of birth. It may mature at any time.
It is likely that the baby’s smile reflex evolved in response to an adaptive challenge: the need to prevent child neglect and infanticide. The preference of adults for smiling faces that is exploited by the infant may well have evolved in contexts other than parental care. We know of other examples where signals evolved to exploit a pre-existing bias in the receiver (RYAN 1998). Since smiling at faces is an effective response to an important adaptive challenge, the infant’s smile reflex qualifies as a special-purpose’sociobiological’ module. Apes have facial emotional expressions related to human smiling (GOODALL 1988, pp273–276), but true human-style smiling took shape only during the past 5 million years, since the human–chimpanzee split, most likely by jumbling up and rearranging elements from older facial-expression circuitry. It remains to be seen whether infant apes direct smiling-related facial expressions at their mothers. P&P seem to imply that the neocortex is too recent to have permitted the evolution of specialized hard-wired circuitry. However, neocortex of sorts has been around for more than 100 million years. If the smile reflex could evolve in a mere 5 million years, then there has been plenty of time for the evolution of hard-wired circuits in the neocortex.
The example of the smile reflex suggests that the best functional models for behavior are frequently those that have been developed by classical ethology over the past century. According to ethologists, the smile reflex is an instinctive response that is triggered by a key stimulus (a face) that activates an internal template (the face cell circuitry) which in turn is connected to a circuitry that patterns the motor output (the ‘smile center’ in the cingulate cortex). Since ‘instinct’ is out of fashion, we may well call the overall circuitry a ‘module’; or we can describe the face cell circuitry and the smile center as two modules arranged in series. In the ethological model the input to the hard-wired circuitry is a sensory stimulus and the output is an observable behavior. In order to accommodate conscious perception, willed action and reasoning, we have to extend this framework by acknowledging that either the output of the circuitry or its input or both may consist of the elements of a cognitive representation. For the time being, this expanded ethological model is more useful than the lofty models of cognitive architecture derived from research in artificial intelligence.
The existence of hard-wired motivational systems at the cortical level is indeed uncertain. Patients with damage to the orbitofrontal cortex show behavioral disinhibition, poor decision making and sociopathy, especially if the damage is acquired early in life (BLAIR/CIPOLOTTI 2000; DAMASIO 1994; DUFFY/CAMP-BELL 1994; PRICE et al. 1990). These patients have normal intelligence, and their working memory seems to be intact. We can speculate that the orbitofrontal cortex analyzes input from the cognitive system and sends inhibitory signals to subcortical emotional and motor systems. The cognitive input would consist of mental representations of intentions, emotions and interpersonal relations in those constellations that would cause damage to self or others if the intentions were acted out. The experience with neurological patients seems to show that proper social conduct and even what appears to be foresight are produced by ‘morality modules’ in the orbitofrontal cortex. These modules may also feed back into the cognitive system as part of the ‘central executive’, to direct the processing of cognitions in working memory. Other modules in the same brain area may function as a ‘parliament of instincts’, coordinating signals from subcortical motivational systems to feed into the central executive of working memory. Other projections may go from the subcortical systems
Evolution and Cognition ❘ 33 ❘ 2001, Vol. 7, No. 1
Gerhard Meisenberg
straight into the buffers of working memory to produce conscious feelings. If all this seems too messy to be true, ask a neuroanatomist to describe the fiber connections in this brain region (PRICE/CARMICHAEL/ DREVETS 1996)! Of course, the extent to which orbitofrontal functions are truly modular or merely modularized, and how modularization is achieved, are wide open questions.
And what about the subcortical systems that P&P champion as the closest approximation to sociobiological modules? My first objection to their description is that it exaggerates the achievements of subcortical neuroscience. True, there is an abundant literature about the brain systems that underlie autonomic regulation, arousal, and even some specific emotional responses, but we still don’t know how it all fits together. For example, electrical self-stimulation of subcortical brain sites has been studied in rats for nearly half a century, since 1953. And yet, we still have no good description of the underlying circuitry. We don’t even know whether self-stimulation sites in places such as the midbrain, lateral hypothalamus and septal area belong to the same reinforcement circuitry or whether they represent functionally independent systems. We don’t even know the exact role of the mesolimbic dopamine system in this circuitry although we have drugs such as amphetamine that can produce euphoria by stimulating this system (KALIVAS/NAKAMURA 1999; ROBBINS/EVERITT 1996; SPANAGEL/WEISS 1999).
There can be no reasonable doubt that the subcortical systems mentioned by P&P are hard-wired. And still, some of them are not good examples for ‘sociobiological’ modules because they did not evolve in their present form in response to a specifiable adaptive challenge. Thus, the cells of the mesolimbic dopamine system become active when the animal either experiences or expects reward, and they reduce their activity when the expectation of reward is frustrated (SCHULTZ 1998). Together with the observation that psychostimulant drugs activate this system, this points to a specific involvement in positive reinforcement and possibly even the conscious experience of pleasure. However, the same neurons are also activated during stress, for example when a rat expects a painful electrical shock or is immobilized on the laboratory bench with duct tape (D’AN-GIO/SERRATO/SCATTON 1990; IMPERATO et al. 1992). Unless laboratory rats are masochists, the dopamine cells are unlikely to function as mediators of pleasure or positive reinforcement in these situations. We can still save the function-specific model by claiming that mesolimbic dopamine cells belong to an incentive system that is recruited whenever an active behavioral response to an environmental stimulus is required. Still, the ‘meaning’ of mesolimbic dopamine activity in different situations may range all the way from ecstasy to panic. It all depends on activities that are going on in other parts of the brain at the same time.
The serotonin system is another example. According to P&P, animals appear to be “relaxed, satisfied and confident” when serotonin is high, but they also mention that knockout mice with markedly reduced serotonin release are less fearful in situations where animals normally exhibit heightened fear responses. The situation is confusing because serotonin release is affected in many other situations as well. It participates in the regulation of REM sleep (MONTI/MONTI 2000), and the effects of fear-inducing and stressful stimuli on the serotonin system are so variable and situation-dependent that any speculation about a unitary function for this neurotransmitter seems hopeless (CHAOULOFF 2000). Serotonin seems to have so many functions that we cannot assign it to a ‘fear module’, a ‘sociality module’, a ‘relaxation module’, or a ‘sleep module’.
Perhaps the closest subcortical approximation to a function-specific module that evolved in response to a specific environmental challenge is the system of conditioned fear described by LEDOUX (LEDOUX 1996). In this case the input is an association between a sensory stimulus and an unpleasant outcome, and there are well-defined, hard-wired outputs into the somatomotor, autonomic, endocrine, and possibly cognitive systems. Presumably this system evolved as a solution to a recurrent adaptive problem: to avoid situations in which unpleasantness has been experienced in the past. Although it accepts cognitive as well as sensory input, it operates automatically and is not easily controlled by the cognitive system, as evidenced by the difficulty of treating phobic patients.
Taken together, the most reasonable model is that the brain does have adaptively meaningful hardwired connections that function in a modular fashion: reflex-like and instinct-like. In typical cases, such stimulus–response arcs have distinct circuitries for sensory analysis, such as the ‘face cells’ in the ventral temporal lobe, and motor circuitries such as the ‘smile center’ in the cingulate cortex. Hard-wired circuitries that mature with minimal need for specific sensory inputs—although they may depend on trophic influences from their afferents—are more typical for subcortical areas but do occur in cortical regions as well, especially in and around the sensory
Evolution and Cognition ❘ 34 ❘ 2001, Vol. 7, No. 1
and motor cortices. We also have domain-general systems that can be recruited by the function-specific modules. These ‘guns for hire’ include, among others, the serotonin system and perhaps the mesolimbic dopamine system at the subcortical level, and the cognitive system at the cortical level.
The relationship between special-purpose modular systems and the cognitive system becomes of paramount importance when the output is not a behavior but a reasoning routine. Thus COSMIDES and TOOBY note that social contract reasoning does not simply reflect a facilitation of the same type of logical reasoning that is applied to other situations (COSMIDES/TOOBY 1992, pp187–193). They propose the existence of two parallel reasoning systems, one for the physical world and the other for social contracts and cheaters. However, rather than postulating two entirely independent systems, it would be more parsimonious to postulate a single cognitive system in which social as well as physical relationships can be represented. One place to look for the difference between deontic and indicative reasoning are the inputs of working memory, which consist entirely of the pre-chewed products of modular systems: sensory cortex for information about the external world, and subcortical circuitry for information about emotional valence and relevance for behavior. If the sensory systems can analyze their inputs for color, face identity, and biological motion, why not for conformity to the stereotype (or archetype, since it presumably develops from pre-wired circuitry in all normal humans) of a social-exchange situation? And if subcortical emotion systems can highlight important sensory stimuli or important elements of cognitive representations—such as the smell of a fast food shack when you are hungry why should they be unable to highlight the cheater part in the mental representation of a social interaction?
Another place to look for specialized deontic circuitry is in the ‘central’ executive of working memory which is, most likely, rather decentralized. We can hypothesize that the executive system contains reasoning modules which use the cognitive representation of the social-contract situation as their input and send their own outputs back into the cognitive system, for example to generate alternative scenarios about possible cheating. The empirical evidence that deontic reasoning differs from reasoning about the physical world in kind and not only in the degree to which general-purpose logic is recruited could simply mean that some components of the central executive respond only to social-contract situations. This would be sufficient to provide a “guidance system” (TOOBY/COSMIDES 1992) for adaptive behavior.
Evolutionary psychologists may question the evolvability of brain systems that are not specifically devoted to the solution of a single adaptive problem. However, guns for hire can evolve from functionspecific systems when elements of these systems are co-opted for new behavioral contexts, either with or without the loss of their original function. Thus, the brainstem reticular formation most likely evolved as a motor center in the first vertebrates for the purpose of coordinating swimming movements and mediating the impact of sensory stimuli on these movements (MASINO 1992). It lost most of its motor functions when motor control shifted to higher centers in midbrain, cerebellum, basal ganglia, and cerebral cortex. Coarse adjustments of muscle tone and the activation of higher brain areas including the cerebral cortex are now its main functions, and it gets recruited whenever these functions have to be adjusted, from sleep–wake regulation to pain responses, orientation to noises, responses to danger, and driving on New York City highways.
The cognitive system, with working memory and its long-term memory system in the hippocampus and adjacent areas of the medial temporal lobe, may have evolved from a short-term memory system that enabled the animal to associate stimuli that were perceived with a time delay. A brain without shortterm memory would not be able to associate stimuli for classical conditioning unless they are received at exactly the same time (CLARK/SQUIRE 1999). Later on, this system was elaborated as a cognitive mapping device to construct a mental model of the environment, for use during foraging and other activities (including taxi driving, MAGUIRE et al. 1997) that required navigation in the environment. Finally, it acquired ever more complex devices to manipulate the mental models in working memory, thus creating the ability to construct models of what could be rather than what is or has been.
These are merely evolutionary just-so stories, but they show that we can build evolutionary scenarios for domain-general multi-function brain systems. The theoretical framework of contemporary evolutionary psychology, with its emphasis on domainspecific, function-specific mechanisms, is too narrow. We have to be aware of the adaptive challenges and selective pressures that shaped our brain, but we also have to be aware of older adaptations that were already in place when a new challenge was encountered. We have to realize that ancient adaptations
Evolution and Cognition ❘ 35 ❘ 2001, Vol. 7, No. 1
can assume new functions and that new functional systems can be created by re-wiring components from older ones.
Evolutionary psychologists will have to accept the existence of multi-purpose ‘guns for hire’ in the brain. They will also have to realize that the constructs of classical ethology are likely to prove more productive as guides for future research than the information-processing metaphors derived from AI. Finally, they have to widen their outlook by dwelling not only on adaptations that evolved, but also on those that could have evolved but didn’t. For example, according to HAMILTON’s rule (HAMILTON 1964) you should love your brother half as much as yourself and your half-brother one quarter as much. And yet, you will never know how much you should love your brother unless you send DNA from yourself, your brother and your putative father to the paternity lab. We never evolved a dependable, hard-wired system for kin recognition. Evolutionary psychologists should point out that this deficiency is central to the evolution of human sociality because it enables us to deploy kin-selected forms of altruism promiscuously.
There are constraints on the number and complexity of genetically hard-wired brain circuits. According to the latest counts, we have only a little more than 30,000 genes in our genome (VENTER et al. 2001). Perhaps we cannot have any more because the rate of detrimental mutations is so high (EYRE-WALKER/KEIGHTLEY 1999; GIANNELLI/GREEN 2000) that we wouldn’t be able to maintain a larger genome. Most of our genes are either housekeeping genes that are expressed in most or all cells, or they support specialized functions of terminally differentiated cells, or they are involved in specialized developmental processes other than brain development such as building the heart, the eye, and the immune system. There can be only a few thousand genes at most that are devoted mainly to the construction of specialized brain circuits, and few if any of them may play roles in the development of one and only one ‘module’. Because of this genetic poverty, combined with the intrinsic mindlessness of the evolutionary process, behavioral adaptations
Even if evolutionary psychologists widen their outlook along these lines, I doubt that a separate discipline named evolutionary psychology can persist. Sociobiology and evolutionary psychology were responses to the biology-neglect of the mainstream psychology and social science traditions of the 20th century. As soon as scholars in the psychological and social sciences incorporate evolutionary thinking as a reality check for their own theories, a separate discipline of evolutionary psychology will no longer be needed. Its historical mission will be fulfilled.
And what can neuroscientists do to approach the mystic unity between their own science and the behavioral sciences? Overspecialization is a far more pressing problem than evolution-neglect. One very specific bug that P&P illustrate only too well, possibly without noticing, is the division of neuroscientists into anthropocentric and ratocentric camps. Most of the neurochemical research about biogenic amines, neuropeptides, and other neurotransmitters and modulators has been done in rats. Also research on subcortical emotional systems is generally done in rats. For lack of human volunteers, anthropocentric neuroscientists do most of their work with monkeys although the advent of functional neuroimaging has permitted complementary studies in humans. Anthropocentric research concentrates on the cortical mechanisms of sensation, motor control, memory and cognition. Subcortical mechanisms are neglected, and the study of emotion in primates is lagging far behind the study of other higher brain functions.
P&P stress, correctly I presume, that species differences tend to be smaller for early-evolved subcortical systems than late-evolved cortical systems. And yet, species differences in social bonding exist even between the closely-related prairie voles and meadow voles that they cite as examples of neurochemically specific social bonding. Monogamous habits in these species are affected by oxytocin, but even if a homologous circuitry exists in humans, chances are it is modulated not by oxytocin but by other neurochemicals. Sure enough, many of the early-evolved systems, such as those concerned with fear responses
must by necessity be crude and imperfect. Because of this imperfection, adaptationist reasoning is only moderately useful for the prediction of cognitive–behavioral mechanisms although it is essential as a reality check for psychological theorizing.
No Evolution. No Cognition
AAK & JULES PANKSEPP postulate a striking dichotomy between ‘genetically dedicated circuits’ for emotions and generalpurpose computational space. The former are phylogenetically ancient subcortical structures, or neurochemical operating systems, which have homologies in many species, and reflect fitness concerns; the latter is subserved by plastic neocortex. The research program of neuroevolutionary psychobiology aims to elucidate the way in which human abilities emerge from developmental interactions between these two mechanisms. The subcortical emotional and motivational systems constitute “the essential character of the human mind”; they may have distinct variants in “different lines of J
Abstract
Although scholars in the natural and human sciences will generally disavow any belief in distinct material and immaterial substances contemporary debates are phrased largely in terms that would have been familiar to the Greek philosophers, and which still divide human characteristics into ‘divine’ or ‘transcendent’ attributes—in modern terminology the surrogate terms are rational, cognitive, discursive, autonomous and creative—and ‘animal’ or ‘corporeal’ attributes the surrogates being emotional, instinctive, determined, immutable, and bounded. This essential dualism preserves the three key dichotomies of mind/ body, cognition/emotion and nature/nurture found in many, if not most, discussions of human nature. Neuroevolutionary psychobiology’s concern to divide the brain into determined affective components and unbounded plastic neocortex by employing such concepts as ‘exaptations’, ‘spandrels’, and ‘emergence’ resides firmly within this quasi-theological Western philosophical tradition. This dualistic approach provides no coherent foundation for the critique of evolutionary psychology.
Key words
Modularity, developmental systems theory, dualism, spandrel, exaptation, brain evolution.
the human family”, may owe their existence to group selection, and can “regulate the construction of personality differences, as well as social systems”. Evolutionary psychologists are exhorted to adopt this model and thereby enter the ranks of the ‘data constrained’ instead of pursuing the ‘tradition’ of COSMIDES and TOOBY, which is described as a “potentially virulent strain of neo-DARWINIAN thinking”.
Although the PANKSEPPs describe ‘seven sins’ these embody four complaints: evolutionary psychologists a). search for adaptive modules where there is only general-purpose computational space; b). are anthropocentric; c). conflate emotions and cognitions, and d). have an anti-organic bias. However, the PANKSEPPs readily acknowledge evidence for the existence of ten or more modules in the neocortex, and evidence for the functional integration of cognition and emotion demonstrated by the work of DAMASIO, LEDOUX, and ROLLS. The PANKSEPPs’ rejection of evolutionary psychology emerges from their belief that neocortical adaptations should be innate, discrete, and detached from subcortical structures, which in turn rests on theoretical misconceptions and their mischaracterization of the neurofoundational issues.
Developmental Systems Theory and Innateness
The PANKSEPPs claim to have assimilated the prescriptions of develop-
mental systems theory, as embodied in the work of OYAMA (2000b) and GRIFFITHS (1997), but throughout their paper they refer to “genetically dedicated circuits” and “genetically dictated adaptations”, which suggests that they are not aware that this theory explicitly opposes the notion of genes as privileged causal entities. Indeed, OYAMA has said that “the idea that traits are ‘transmitted’ in heredity, rests on notions of genetic programming that are ultimately quite preformationist” (OYAMA 2000a, p21). Following LEHRMAN’s (1953) lead, developmental systems theorists argue that nature selects for outcomes. Any of the inherited components of the developmental system, which includes genes,
Evolution and Cognition ❘ 39 ❘ 2001, Vol. 7, No. 1
Ian Pitchford
other biological resources, patterns of maternal care, play, and so forth can ‘mutate’ producing novel phenotypic characteristics. In contrast to the perspective encouraged by the ‘genetic blueprint’ or ‘genetic program’ metaphors, this approach allows for phenotypic variability, and for constant, stable outcomes, provided that all of the resources required by the developmental system are available (GRIFFITHS 1997, p186). This theory is applicable to the whole of evolutionary biology, and not only to the neocortex as the PANKSEPPs seem to think.
Perhaps the most insidious consequence of the genetic blueprint idea is the expectation that phenotypic characteristics are innate, meaning ‘hereditarily determined’, or arising independently of environment or experience (LEHRMAN 1953). Describing a trait as ‘innate’ confuses at least four properties that can vary independently: “(1) that it is found in an individual because of their ancestry rather than their current environment; (2) that its growth does not depend on that environment for anything but basic sustenance; (3) that it is present at birth or early in development; and (4) that it is part of the ‘nature’ of the species… The result of this mismatch between concept and reality is that when theorists discover that one element of the innateness concept applies to a trait, they are liable to assume that the other elements must also apply” (GRIFFITHS 1997, p104). Our faculties are the product of developmental systems, and consequently they are neither innate nor structured by the environment. The PANKSEPPs formulation perpetuates the very ‘nature versus nurture’ dichotomy that developmental systems theory aims to transcend.
Adaptations, Exaptations, and Spandrels
The PANKSEPPs are surely correct that we should be confident where convergent lines of evidence point in the same direction, which makes their assertion that recent brain evolution is characterised by “the rapid expansion of general-purpose cortico-computational space (which permitted the emergence of foresight, hindsight and language)” appear strikingly incongruous. The obfuscations of those who like to characterise themselves as opposed to ‘DAR-WINIAN fundamentalism’ (GOULD 1997) play a role here. The PANKSEPPs tell us that “since the emergence of massive, general-purpose cortical space, exaptations and spandrels have arisen everywhere we look”. Their mentor GOULD (1984) does not disagree with biology’s emphasis on natural selection but believes “that we have become overzealous about the
power and range of selection by trying to attribute every significant form and behavior to its direct action”. Obviously, we should not be interested in ‘attributing’ anything at all to natural selection. We need to look at the evidence that “a function is served with sufficient precision, economy, efficiency, etc. to rule out pure chance as an explanation” (WILLIAMS 1996, p10).
In their original paper on ‘spandrels’ GOULD/ LEWONTIN (1979) make entirely prosaic observations concerning the ubiquity of phyletic constraints, and argue that the evidence for Aztec cannibalism, the chin, and papillary ridges as adaptations is not strong. In a second paper cited by the PANKSEPPs GOULD (1991) observes that ‘exaptations’ can be defined as characters not selected for their current function which may or may not have arisen originally by the direct action of natural selection. Useful characters that did not arise by the action of natural selection are a type of exaptation, or coopted nonaptation, called a spandrel. GOULD mentions weight, the delayed ossification of skull bones, and language as probable exaptations.
It seems clear from these examples that (with the exception of language, which appears to share little in common with the other examples) spandrels are trivial features that would not be identified as adaptations by anyone using the conventional criteria. It is equally clear that we have no good reason to restrict the term ‘adaptation’ to only those characters that perform the function for which they were first selected. As GRIFFITHS and STERELNY point out “GOULD and VRBA think that a trait is an adaptation only for the purpose for which it was first selected. But what justifies this special status for the first of many selection pressures? The importance of the concept of adaptation in biology is that it explains the existence of many traits of the organisms we see around us. This explanation is not just a matter of how traits first arose, but of why they persisted and why they are still here today” (STERELNY/GRIFFITHS 1999, p219). The only complex functional characteristic claimed as an exaptation is language, and this is done by argument from authority—the authority in question being CHOMSKY, who is said to have “long advocated a position corresponding to the claim that language is an exaptation of brain structure” (GOULD 1991, p61). However, CHOMSKY has not “expressed views on the lack of a role for natural selection in… the origin of language”. On the contrary he believes “that natural selection is operative in this case” (personal communication 1999).
Evolution and Cognition ❘ 40 ❘ 2001, Vol. 7, No. 1
The PANKSEPPs are ambivalent here and acknowledge that “there is substantial neuro-evolutionary evidence for the emergence of special abilities such as language”. Their rejection of it as an adaptation seems to be based in part on the misconception acquired from the work of GOULD and colleagues that only something arising from “de novo genetic shaping” can be characterised as such. It is notable that connectionist models most in keeping with the general-purpose architecture that the PANKSEPPs propose have produced results incompatible with what we know of language processing, acquisition, and pathology. CHOMSKY observed recently that “in the case of language, the evidence for connectionist models is… about zero” (quoted in SMITH 1999, p135). The case of Christopher, a severely retarded autistic savant who remains unable to take care of himself, but who has mastered over twenty languages, seems particularly problematic for the model proposed by the PANKSEPPs. Despite his abilities Christopher was unable to master the artificial language Epun, which violates the rules of natural language. Surprisingly, a control group of bright undergraduates fared no better (SMITH/TSIMPLI 1995). Even if we were to ignore arbitrarily the other evidence for the modularity and evolution of language it makes no sense to interpret these findings in terms of the properties of plastic neocortex. The claim that “language emerged within a spandrel of evolving multimodal brain complexities” is mere sophistry. Instead of relying on the quaint notion that their hypothesis should be taken seriously because it has not been falsified (just like most other hypotheses) the PANKSEPPs should provide what they require of evolutionary psychologists: a plausible account of the distal and proximal mechanisms responsible for generating the complex functionality and patterns of acquisition and dissociation that we observe. To call on ‘learning’ will not suffice. Induction and explanation in science require reference to causal homeostatic mechanisms (GRIFFITHS 1997, pp187– 201), not appeals to folk psychological concepts that do not represent projectable categories (see GRIF-FITHS 1997, pp175–176, and TOOBY/COSMIDES 1992, pp111–118, for useful discussions).
Evolutionary Psychology, Selfishness, and Inclusive Fitness
The PANKSEPPs regard the theory of inclusive fitness to be “most clearly applicable to sub-human species”, and their antipathy to its application to Homo sapiens (or at least to the neocortex) seems to stem from their belief that selfish gene theory implies that all human behaviour is genetically constrained to be selfish. They tell us that “perhaps all too many individuals utilize those capacities largely for their own selfish ends, but it is important to emphasize that the nature of the higher regulatory systems in humans does permit many alternative courses of action”. This misconception appears to underpin their endorsement of group selection. I wont discuss this simple misinterpretation here as it has been dealt with many times in the literature, not least by DAWK-INS (1981, 1989). The improbability (though not impossibility) of group selection as an explanation for evolved traits is a standard text book matter (MA-JERUS/AMOS/HURST 1996, pp110–115), and John MAYNARD SMITH (1998) has described much of the controversy as “largely semantic”.
Unlike sociobiologists, evolutionary psychologists are skeptical of the extent to which evolved behavioural tendencies cause human behaviour to assume the form that maximizes inclusive fitness (SYMONS 1989). BARKOW (1990) has described three ways in which sociocultural traits can affect the fitness of their participants: “a) they can enhance fitness because the cultural trait is a direct reflection of an evolved psychological mechanism; b) they can lower fitness or be neutral for it, because cultural processes are semi-independent of biological evolution; or c) they can enhance fitness epiphenomenally, that is, in a manner having little or no connection with past genetic selection”. Because our contemporary environments (and hence our developmental systems) incorporate such novel aspects as mass media, drugs, processed foods, pollutants, large group sizes, reduced interaction with kin, and many other phenomena not typical of the hunter– gatherer environment, it is highly probable that current behaviour is unlikely to be a reliable guide to ancestral behaviour, and evolutionary psychology’s commitment to the study of evolved psychological mechanisms should take precedence over (though not entirely replace) the study of current fitness.
Neuro-Foundational Issues
For the PANKSEPPs the neurofoundational issues we need to take into consideration are the existence of evolutionary layers in the brain; the preservation of ancient motivational mechanisms; the evidence for ‘mind’ in the variability of behaviour related to intrinsic representations rather than external stimuli; the supervenience of mind on the brain; the irreducibility of mind to the functions of the brain, and an
Evolution and Cognition ❘ 41 ❘ 2001, Vol. 7, No. 1
appreciation that the foundations of human nature were ‘solidified’ way before the Pleistocene era. I would like to suggest a broader perspective based on empirical and theoretical considerations.
In 1935 William KING GREGORY suggested that replication of body parts due to genetic mutation had been a major mechanism in evolution. In 1971 this idea was extended by John ALLMAN’s and Jon KAAS’ proposal that the evolution of the brain was characterised by the replication of cortical areas (AL-LMAN 1999, p40). RILLING’s and INSEL’s comparative MRI study of the primate neocortex confirms the finding that the human brain is slightly over three times larger than would be expected for a primate of the same body size (1999b). However, the data indicate a striking discrepancy between human and pongid brains in the extensive gyrification in the prefrontal cortex of the former, an important finding given the role of this region in complex problem-solving (KOECHLIN et al. 1999), and social intelligence (ROWE et al. 2001; SHALLICE 2001; STUSS/ GALLUP/ALEXANDER 2001). As RILLING/INSEL (1999b) conclude this departure from allometric trends “suggests selection for increased gyrification in the prefrontal cortex throughout hominid evolution”. The other area noted for significantly more gyrification than expected is the seventh coronal slice, a region incorporating WERNICKE’s area, long implicated in the production and comprehension of language (1999b). RILLING and INSEL also note that the increase in human neocortical gray matter is not proportional with the increase in the volume of the rest of the brain and that, although the increase in white matter outpaces that in grey, this increase falls well short of that necessary to retain the same level of interconnectivity between neurons. Another scan of 11 primate species concentrating on the corpus callosum and anterior commissure demonstrates that the increase in primate brain size has resulted in increasingly independent hemispheres (RILLING/INSEL 1999a). Through their work on the insular cortex of bottlenose dolphins MANGER et al. (1998) have found that although brain sizes vary dramatically across animal species, the range of module size is restricted, though the number of cortical areas across species is highly variable (KAAS 1993; KAAS/REINER 1999). For additional evidence on mosaic brain evolution see BARTON/HARVEY (2000) and DE WINTER/OXNARD (2001). These findings confirm that brain evolution is characterised by the independent evolution of brain structures with anatomical and functional links. Indeed, one of the most distinctive features of the neocortex is
its modular organization (JONES 2000; MOUNTCASTLE 1997; ROCKLAND 1998).
The existence of a neuronal type found only in the brains of pongids and hominids is also likely to be of importance. Using samples of the anterior cingulate cortex (BRODMANN’s area 24) of 28 primate species NIMCHINSKY et al. (1999) found a spindleshaped cell in layer Vb specific to humans and great apes. The anterior cingulate is known to be involved in response selection (AWH/GEHRING 1999; TURKEN/ SWICK 1999), and performance monitoring (CARTER et al. 1998), but also appears to have a number of discrete, functional regions subserving important aspects of cognition, emotion, and notably vocalization (BUSH/LUU/POSNER 2000). NIMCHINSKY et al. (1999) note that “the emergence of this unique neuronal type in a neocortical area involved in vocalization in primates coincides with the evolution as a definable anatomic structure of the planum temporale, a region that is important for language comprehension. In view of the language comprehension abilities of great apes, it is therefore possible that several cortical structures involved in the production of specific vocalizations and in communicative skills sustained simultaneous, considerable, adaptive modifications during brain evolution in hominoids”.
In considering neuroevolutionary matters we should always keep the issue of sexual dimorphism in mind. There are two types of human brain, male and female, and it is reasonable to expect that these have been subject to different selection pressures. For example, women have a higher proportion of grey matter to cranial volume, whereas men have a higher proportion of white matter and cerebrospinal fluid to cranial volume. Women also have a relatively larger corpus callosum than men. GUR et al. (1999) found that of the top ten performers in a spatial task, nine were men, and seven of these men had greater white matter volumes than any of the women in the study. Our large brains may not simply provide an excess of plastic neurons capable of subserving any function, but may be a solution to the problem of retaining adequate functioning over a prolonged life span (HUMPHREY 1999), something that could be of particular importance to caregivers. ALLMAN has found that there is a significant correlation between brain weight and maximum life-span in haplorhine primates (ALLMAN/MCLAUGHLIN/ HAKEEM 1993), and that the maximum human lifespan is close to what would be expected for a primate of our relative brain size (ALLMAN 1999, p172). ALL-MAN et al. (1998) have also discovered in a variety of
Evolution and Cognition ❘ 42 ❘ 2001, Vol. 7, No. 1
species that caregivers live longer, whether male or female, and “that there is no difference in survival between the sexes in species in which both parents participate about equally in infant care”. The fact that human females are the primary caregivers, and that human grandmothers are able to enhance their fitness post-menopausally by assisting the reproductive success of their daughters may also help to explain the structural and functional differences between the brains of men and women (HAWKES et al. 1998; O’CONNELL et al. 1999).
All adaptations have costs and benefits, and it is certain that psychological mechanisms are not costfree because the rate of DNA damage in mammalian cells is extremely high, amounting to tens of thousands of DNA damages per day. This implies an enormous metabolic cost in maintenance and repair (DU-KAS 1999). Also, as many of the processes within the brain are mediated by the same neurochemicals, functional systems must have the capacity to ensure that the correct information is elicited as required. One benefit derived from the piecemeal addition of overlapping systems is explained by DUKAS in his analysis of the costs of memory: redundancy helps to reduce the amount of error and noise in the system, and therefore “probably plays a key role in ensuring a high level of accuracy” (DUKAS 1999). The cost of redundancy is in terms of increased brain mass and energetic expenditure on maintenance, repair and replication. As HENINGER (1999, pp93–94) explains: “One of the main features of the nervous system is the mutually dependent, diffuse, and often redundant biologic processes that subserve functions. In contrast to the relative specificity of sensory and motor systems, the systems subserving sleep– wakefulness, arousal-motivation, emotional reactivity, memory, and higher order behavioural functions are more widely distributed anatomically. The systems demonstrate extremely complex nonlinear response characteristics so that there is not a simple one–to–one correspondence between measures of neuronal function and the behaviours studied. In addition, there is a great deal of plasticity so that remaining systems can compensate for deficits”.
The PANKSEPPs are surely right when they claim that ancient phylogenetic mechanisms are conserved, and that consequently homologies can teach us much about the construction of the human mind, but I can think of no reason for believing that the neocortex has been moulded by processes substantially different to those responsible for the subcortical structures involved in the basic emotions. The PANKSEPPs vision of a brain consisting of horizontal layers, derived from MACLEAN’s model of the triune brain, is deeply misleading. Instead of horizontal layers our model should postulate integrated vertical modules built from co-evolving structures distributed throughout the brain, the different subcomponents of which may be traced to different evolutionary eras, and explained by the influence of different selection pressures.
Finally, we should be aware that the distinctive cultural traits of human beings appear to have emerged (or grown in significance) during a period in which brain sizes have decreased. It appears that since the Late Pleistocene (around 30,000 years ago) human brain size has decreased by approximately ten per cent with this decrease being paralleled by a decrease in body size. HENNEBERG (1998) notes “it may be concluded that the gross anatomy of the hominid brain is not related to its functional capabilities. The large human brain:body size ratio may be a result of the structural reduction of the size of the gastrointestinal tract and, consequently, its musculoskeletal supports. It is related to richer, meatbased diets and extra-oral food processing rather than the exceptional increase in the size of the cerebrum. The exceptional mental abilities of humans may be a result of functional rather than anatomical evolution”.
All of the foregoing theoretical and empirical considerations indicate that the sudden emergence of a highly plastic general-purpose neocortex responsible for multimodal functioning is a distinctly implausible evolutionary event, and that evolutionary psychology’s commitment to modularity is sound.
Conclusion
Evolutionary psychologists, including the PANK-SEPPs’ exemplars COSMIDES and TOOBY, advocate the assimilation of findings from evolutionary biology and its related disciplines (TOOBY/COSMIDES 1998), including neurobiology (TOOBY/COSMIDES 1995), in a vertically integrated hierarchy of mutually consistent scientific disciplines that does not require or call “for reductionism nor for the conquest and assimilation of one field by another” (COSMIDES/TOOBY/ BARKOW 1992, p12). Evolutionary psychology’s adoption of the concept of modularity from neurology creates a natural affinity with the neurosciences, whilst the emphasis on psychological mechanisms and the mismatch between ancestral and contemporary environments provides a bridge to psychology and the social sciences. This richly interdisciplinary field has already generated an abundance of research
Evolution and Cognition ❘ 43 ❘ 2001, Vol. 7, No. 1
in developmental psychology, neurology, psychiatry, and primatology, particularly on theory of mind, which has produced compelling new theories of autism and schizophrenia. This multi- and interdisciplinary research exhibits the consilience, or confluence of different lines of evidence, so typical
of robust scientific work (see CORCORAN 2000 and BARON-COHEN 2000 for a summary). It compares extremely favourably with that based on the neurochemical individuation of traits and disorders admired by the PANKSEPPs, which has produced so many unsatisfactory theories and treatments (VALENSTEIN 1998).
Neuroevolutionary psychobiology’s reassignment of the tabula rasa to the neocortex revivifies CARTESIAN dualism and represents a desperate strategy likely to be extremely deleterious to scientific
References
- Allman, J. M./McLaughlin, T./Hakeem, A. (1993) Brain weight and life-span in primate species. Proceedings of the National Academy of Sciences USA 90: 118–22.
- Allman, J. M. (1999) Evolving brains. Scientific American Library: New York.
- Allman, J. M./Rosin, A./Kumar, R./Hasenstaub, A. (1998) Parenting and survival in anthropoid primates: Caretakers live longer. Proceedings of the National Academy of Sciences USA 95: 6866–6869.
- Awh, E./Gehring, W. J. (1999) The anterior cingulate cortex lends a hand in response selection. Nature Neuroscience 2: 853–854.
- Barkow, J. H. (1990) Beyond the DP/DSS controversy. Ethology & Sociobiology 11: 341–351.
- Baron-Cohen, S. (2000) Theory of mind and autism: A fifteen year review. In: Baron-Cohen, S./Tager-Flusberg, H./Cohen, D. J. (eds) Understanding other minds: Perspectives from developmental cognitive neuroscience. Oxford University Press: Oxford, pp. 3–20.
- Barton, R. A./Harvey, P. H. (2000) Mosaic evolution of brain structure in mammals. Nature 405:1055–1058.
- Bush, G./Luu, P./Posner, M. I. (2000) Cognitive and emotional influences in anterior cingulate cortex. Trends in Cognitive Sciences 4: 215–22.
- Carter, C. S./Braver, T. S./Barch, D./Botvinick, M. M./Noll, D./Cohen, D. J. (1998) Anterior cingulate cortex, error detection, and the online monitoring of performance. Science 280: 747–749.
- Corcoran, R. (2000) Theory of mind on other clinical conditions: Is a selective ‘theory of mind’ deficit exclusive to autism? In: Baron-Cohen, S./Tager-Flusberg, H./Cohen, D. J. (eds) Understanding Other Minds. Oxford University Press: Oxford, pp. 391–421.
- Cosmides, L./Tooby, J./Barkow, J. H. (1992) Evolutionary psychology and conceptual integration. In: Barkow, J. H./ Cosmides, L./Tooby, J. (eds) The Adapted Mind: Evolutionary psychology and the generation of culture. Oxford University Press: New York, pp. 3–15.
- Dawkins, R. (1981) In defence of selfish genes. Philosophy 56: 556–73.
- Dawkins, R. (1989) The selfish gene. Oxford University Press: Oxford.
- de Winter, W./Oxnard, C. E. (2001) Evolutionary radiations and convergences in the structural organization of mammalian brains. Nature 409: 710–714.
- Dukas, R. (1999) Costs of memory: Ideas and predictions. Journal of Theoretical Biology 197: 41–50.
- Gould, S. J. (1984) Only his wings remained. Natural History 93: 10–18.
- Gould, S. J. (1991) Exaptation: a crucial tool for an evolutionary psychology. Journal of Social Issues 47: 43–65.
- Gould, S. J. (1997) Darwinian fundamentalism. New York Review of Books 44: 34–37.
- Gould, S. J./Lewontin, R. C. (1979) The spandrels of San Marco and the Panglossian paradigm: a critique of the adaptationist programme. Proceedings of the Royal Society of London B 205: 581–598.
- Griffiths, P. E. (1997) What emotions really are. University of Chicago Press: Chicago IL, London.
- Gur, R. C./Turetsky, B. I./Matsui, M./Yan, M./Bilker, W./ Hughett, P./Gur, R. E. (1999) Sex differences in brain gray and white matter in healthy young adults: correlations with cognitive performance. Journal of Neuroscience 19: 4065–4072.
- Hawkes, K./O’Connell, J. F./Blurton Jones, N. G./Alvarez, H/ Charnov, E. L. (1998) Grandmothering, menopause, and the evolution of human life histories. Proceedings of the National Academy of Sciences USA 95: 1336–1339.
- Heninger, G. R. (1999) Special challenges in the investigation of the neurobiology of mental illness. In: Charney, D. S./ Nestler, E. J./Bunney, B. S. (eds) Neurobiology of Mental Illness. Oxford University Press: Oxford, pp. 89–99.
- Henneberg, M. (1998) Evolution of the human brain: Is bigger better? Clinical & Experimental Pharmacology & Physiology 25: 745–749.
- Humphrey, N. (1999) Why human grandmothers may need large brains. Psycoloquy 10(024).
- Jones, E. G. (2000) Microcolumns in the cerebral cortex. Proceedings of the National Academy of Sciences of the USA 97: 5019–5021.
- Kaas, J. H. (1993) Evolution of multiple areas and modules within neocortex. Perspectives in Developmental Neurobiology 1:101–107.
- Kaas, J. H./Reiner, A. (1999) The neocortex comes together. Nature 399: 418–419.
- Koechlin, E./Basso, G./Pietrini, P./Panzer, S./Grafman, J.
Evolution and Cognition ❘ 44 ❘ 2001, Vol. 7, No. 1
(1999) The role of the anterior prefrontal cortex in human cognition. Nature 399: 148–151.
- Lehrman, D. S. (1953) A critique of Konrad Lorenz’s theory of instinctive behavior. Quarterly Review of Biology 28:337– 363.
- Majerus, M./Amos, W./Hurst, G. (1996) Evolution. Longman: London.
- Manger, P./Sum, M/Szymanski, M/Ridgway, S. H./Krubitzer, L. (1998) Modular subdivisions of dolphin insular cortex: Does evolutionary history repeat itself? Journal of Cognitive Neuroscience 10:153–166.
- Maynard Smith, J. (1998) The origin of altruism. Nature 393: 639–40.
- Mountcastle, V. B. (1997) The columnar organization of the neocortex. Brain 120: 701–722.
- Nimchinsky, E. A./Gilissen, E./Allman, J. M./Perl, D. P./Erwin, J. M./Hof, P. R. (1999) A neuronal morphologic type unique to humans and great apes. Proceedings of the National Academy of Sciences USA 96: 5268–5273.
- O’Connell, J. F./Hawkes, K./Blurton Jones, N. G. (1999) Grandmothering and the evolution of Homo erectus. Journal of Human Evolution 36: 461–485.
- Oyama, S. (2000a) Evolution’s Eye. Duke University Press: Durham NC.
- Oyama, S. (2000b) The Ontogeny of Information. 2nd edition. Duke University Press: Durham NC.
- Rilling, J. K./Insel, T. R. (1999a) Differential expansion of neural projection systems in primate brain evolution. Neuroreport 10: 1453–1459.
- Rilling, J. K./Insel, T. R. (1999b) The primate neocortex in comparative perspective using magnetic resonance imaging. Journal of Human Evolution 37:191–223.
- Rockland, K. S. (1998) Complex microstructures of sensory cortical connections. Current Opinion in Neurobiology 8: 545–551.
- Rowe, A. D./Bullock, P. R./Polkey, C. E./Morris, R. G. (2001) ‘Theory of mind’ impairments and their relationship to executive functioning following frontal lobe excisions. Brain 124: 600–616.
- Shallice, T. (2001) ‘Theory of mind’ and the prefrontal cortex. Brain 124: 247–248.
- Smith, N. V. (1999) Chomsky: Ideas and ideals. Cambridge University Press: Cambridge.
- Smith, N. V./Tsimpli, I.-M (1995) The mind of a savant: Language learning and modularity. Blackwell: Oxford.
- Sterelny, K./Griffiths, P. E. (1999) Sex and death. University of Chicago Press: London.
- Stuss, D. T./Gallup, G. G./Alexander, M. P. (2001) The frontal lobes are necessary for ‘theory of mind’. Brain 124: 279– 86.
- Symons, D. (1989) A critique of Darwinian anthropology. Ethology & Sociobiology 10: 131–144.
- Tooby, J./Cosmides, L. (1992) The psychological foundations of culture. In: Barkow, J. H./Cosmides, L./Tooby, J. (eds) The Adapted Mind: Evolutionary psychology and the generation of culture. Oxford University Press: New York, pp. 19–136.
- Tooby, J./Cosmides, L. (1995) Foreword to Mindblindness by Simon Baron-Cohen. MIT Press: Cambridge MA, pp. xi– xviii.
- Tooby, J./Cosmides, L. (1998) Evolutionizing the cognitive sciences: A reply to Shapiro and Epstein. Mind & Language 13: 195–204.
- Turken, A. U./Swick, D. (1999) Response selection in the human anterior cingulate cortex. Nature Neuroscience 2: 920–924.
- Valenstein, E. S. (1998) Blaming the brain The Free Press: New York.
- Williams, G. C. (1996) Adaptation and natural selection. Princeton University Press: Princeton NJ. Originally published in 1966.
The Case for Modularity: Sin or Salvation?
Introduction
The central thesis of PANKSEPP and PANKSEPP (2001, henceforth, P&P) is that there are no specially evolved, higher-order domain-specific cognitive mechanisms to speak of. Apart from the basic DARWINIAN emotions (fear, surprise, anger, sadness, joy, disgust), so-called cognitive ‘modules’ are more likely “the product of dubious human ambition rather than sound scientific reasoning”. What evolutionary psychologists (as well as many cognitive and developmental psychologists) take to be modular structures are actually the epigenetic products of the ancestral emotional functions of the brain (rooted in what was once called “the limbic system,” MACLEAN 1990) and more recent “general-purpose brain mech-
Abstract
The case for evolved cognitive modules rests on several converging lines of evidence: Functional design (analogy), ethology (homology), universality, precocity of acquisition, independence from perceptual experience (poverty of stimulus), selective pathology (cerebral impairment), resistance to inhibition (hyperactivity), ease of cultural transmission. No factor may be necessary but evidence for all or some is compelling, if not conclusive, for domains like folkmechanics, folkbiology, folkpsychology. By constrast, no empirical evidence supports PANKSEPP and PANKSEPP’s central thesis: that what evolutionary psychologists (and many cognitive and developmental psychologists) consider modular structures are actually epigenetic products of subcortical mammalian emotions and neocortical general-purpose computations. Arguably, no significant empirical discovery about language or other higher-order cognitive structures yet owes to inquiries about evolutionary origins and functions. Nevertheless, adopting evolutionary psychology’s requirement that candidate exaptations and spandrels be described, as far as possible, in connection with evolved adaptations, opens new avenues for exploring and testing modular designs.
Key words
Modularity, domain-specificity, language, folkpsychology, folkbiology.
anisms”. In more than a dozen places, P&P repeat as mantra that empirical evidence indicates the modern human mind was created through the dual functioning of subcortical mammalian emotions and a neocortical general-purpose computational device. Furthermore, this highly flexible, all-purpose intelligence probably emerged “more rapidly via group selection than by individual selection”.
There is much to commend the cautionary tale that P&P tell regarding possible excesses of sociobiological speculation, including what are arguably ‘just-so’ stories about modularized adaptations for
rape (THORNHILL/PALMER 2000), homicide (BUSS 1999) and emotional disorders such as depression (NESSE/LLOYD 1992). P&P’s position on the derivative and epigenetic character of ‘secondary’ or ‘social’ emotions (guilt, love, empathy, etc.), is also defensible (e.g., DAM-ASIO 1994), although seriously debatable (cf. LEDOUX 1996; GRIFFITHS 1997). Finally, P&P’s take on the current, overlyfunded fad for neuroimaging (driven more by technological innovation and the industry it supports than by any theoretical insight) is credible. There can no more be a theory directly derived from observations of neuroimaging than there can be a theory derived directly from observations of meter readings; it is only in the service of some prior abstract theory that such observa-
tions can make sense. Nevertheless, there is no empirical evidence whatever to indicate that P&P’s central thesis is true, and much to suggest it is false.
Evolutionary Psychology: No Explanatory Value for Higher-Order Cognition (Yet)
To-date, evolutionary psychology has not predicted or discovered any significant or surprising aspect of language or higher-order cognitive devices of the sort I describe below. But this does not mean that all
Evolution and Cognition ❘ 46 ❘ 2001, Vol. 7, No. 1
evolutionary psychology accounts are ‘just-so’ stories. The best account of language as an evolutionary adaptation centers on the claim that “language shows signs of design for the communication of propositional structures over a serial channel” (PINKER/BLOOM 1990). PINKER and BLOOM describe how specific syntactic structures conform to this language-specific design in ways that provide functional advantage: for example, through certain structure-dependent rules (phrase structure) and principles of embedding (recursion) that allow the formulation and expression of infinitely many discrete ideas by finite and few means. This enables the multiple thoughts of multiple individuals to be combined, tested in imagination and consequently included or excluded from having a role in some future action. A population whose individuals could contemplate alternative scenarios in any sequence and at any rate, benefit from the cognitive travails of others, and let conjectures die instead of themselves, would surely have had an evolutionary advantage over a population that couldn’t.
PINKER and BLOOM offer a well-reasoned and often nonobvious analysis of evolutionary tradeoffs involved in opting for linear communication through an auditory medium: visual displays better communicate highly complex topological relationships (“a picture is worth a thousand words”), gestures better convey emotion, and language is inefficient transmitting information about taste or smell. For all its insight and plausibility, though, PINKER and BLOOM’s account remains wholly backward-looking: no structural discovery, novel prediction or theoretical breakthrough ensues within the CHOMSKYEAN framework that the authors adopt.
According to Jerry FODOR (1998), whose reasoning is close to CHOMSKY’s (1988, p170), it is not likely that natural selection gradually produced an adaptive mutation for a language instinct. Rather, selection pressures unrelated to language simply made human brains a little larger and more complex. Unlike gradual and incremental adaptations, a little added complexity can go a long way fast to produce multiple novel structures:
“Make the giraffe’s neck just a little bit longer and you correspondingly increase, just by a little, the animal’s capacity to reach the fruit at the top of the tree… But make an ape’s brain just a little bigger (or denser, or more folded, or, who knows, grayer) and it’s anybody’s guess what happens to the creature’s behavioural repertoire”. (FODOR 1998).
Much ink has spilled and acrimony vetted in the debate between evolutionary psychologists and the scientists who criticize them. In the case of language, however, no empirical issue has yet to turn on the debate. Despite very different evolutionary stories, CHOMSKY, FODOR, PINKER and BLOOM fundamentally agree on the specific computational structures that characterize language, on its innateness, on its highly specialized mode of operation (modularity) and on the fact that all of this is uniquely the product of evolution, whether adaptation or by-product.
The Language Module
From a purely logical standpoint, if a mind is able to take fragmentary instances of experience (relative to the richness and complexity of the whole data set) and spontaneously predict (project, generalize) the extension of those scattered cases to an indefinitely large class of intricately related cases, then the inferential structure responsible for this prediction cannot possibly derive from the experience. As Hume noted, the structure must be prior to experience, just like the cranes and architects used for constructing buildings must exist prior to initial construction: a building does not just build itself. There is no other possibility. What, then, are the physical possibilities for a mental structure to be “prior to experience”?
One could hold with SOCRATES or BUDDHA that people are born with a past-life ‘memory’ for the structure of experience they will encounter later in life; or, one could hold with Thomas AQUINAS or Immanuel KANT that God put structure in people’s minds. If, however, one accepts humans as biological creatures whose species attributes emerged through the same evolutionary processes that govern the emergence of all other species, then there appears to be no alternative to a priori mental structures being evolved biological structures (whether as adaptations or by-products of adaptations). To say an evolved biological structure is ‘innate’ is not to say that every important aspect of its phenotypic expression is ‘genetically determined’. Biologically poised structures channel development, but do not determine it—like mountains that channel scattered rain into the same mountain–valley river basin (WADDINGTON 1959).
P&P might grant all this, but argue that an allpurpose, domain-general computational device suffices to give the mind/brain inferential power beyond the information given. As an empirical claim, this entails that one should be able to deduce the structural principles and parameters governing any specific domain from general purpose-structures acting under the constraints of experience in the rele-
Evolution and Cognition ❘ 47 ❘ 2001, Vol. 7, No. 1
vant domain. For example, P&P offer that language “may be closely linked to anterior cingulate and adjacent frontal lobe tissues which appear to mediate certain types of pain, feelings of separation distress and thereby social sensitivities” (cf. MACLEAN 1990). But there is nothing in what we know of general intelligence (e.g., conditioning, association, etc.), pain, distress, social sensitivity, or the structures of anterior cingulate and adjacent frontal lobe tissues, to even remotely hint at the highly structured, and structurally peculiar, principles and parameters of human language. This includes: anaphora (the structural constraints on how even widely separated parts of an expression co-refer), quantification (the structural constraints that ‘who,’ ‘many,’ few,’ etc. impose on the syntactic behavior of subjects and objects), negation (it is literally impossible to learn negation through perceived experience or ‘interaction,’ as one cannot perceive or interact with something that isn’t), word order (all languages have a specified linear order linking arguments to predicates), and so forth (CHOMSKY 1982).
Briefly, the central tenet of CHOMSKY’s (2000) approach is that there is a language system, LS, of the human brain. LS reflexively discriminates and categorizes parts of the flux of human experience as ‘language,’ and develops complex abilities to infer and interpret this highly structured, and structurally peculiar, type of human production. In a general sense, there is nothing intrinsically different about LS—in terms of innateness, evolution or universality—than the visual system (VS), immune system (IS), respiratory system (RS), or any other complex biological system. Learning syntactic structure through ‘social interaction’ is no more plausible an alternative than learning by ‘osmosis’. If a child is initially deprived of linguistic input, the child may never come to acquire, or ‘know,’ much in the way of language, just as a kitten initially deprived of normal exposure to the visual world may never come to ‘know’ much by way of object recognition (HUBEL 1988). Still, emotion and social interaction no more ‘create’ language in the child’s mind than lighting and object patterning ‘create’ vision in the kitten.
Interacting Modules
LS is no more (or less) ‘autonomous’ from the surrounding social environment, or from other mental systems, than VS is detachable from surrounding light and object patterning or from other physical systems (including, in humans, linguistic and other cognitive systems of meaning, MARR 1982). LS and VS do not exist, and cannot develop, in isolation, but only as subsystems of even more intricate structures. Thus, claims about the biological ‘autonomy’ or ‘modularity’ of LS or VS refer only to a specifiable level of systemic functioning within a system hierarchy. A difficult empirical issue concerns the extent to which other cognitive ‘performance’ systems are themselves specifically adapted for language. There is little doubt that the sound system is highly structured for access (psycholinguistic experiments with neonates and even fetuses, dichotic listening experiments, comparative phonology, and so forth are compelling). The interface between syntactic and semantic systems is much more obscure. Speculation is rampant, debate is furious, and critical experiments are few, as might be expected from a relatively new and dynamically changing science. But progress is being made, slowly and laboriously, as intense research by teams of well-trained investigators continues.
Cognitive and developmental psychologists and psycholinguistics have identified a number of structural principles in human cognitive systems that relate to the interface between LS and these other systems. Among the principles discovered is “the whole–object constraint” (CAREY 1985). Children, whatever their culture or language, apparently assume as a default that nouns in general apply to whole objects (e.g., a rabbit) and not to parts of the object (e.g., a piece of a leg, a patch of fur, or spatially separated patches of fur and pieces of leg), or the object–and–its–environment. Children actually have to learn that this is not always the case (just as children have to learn that the shadow of an object is not itself an object, SPELKE et al. 1995). This apparently ‘innate bias’ helps to solve the problem of radical indeterminacy of translation. Consider an anthropologist who visits an exotic tribe for the first time and sees a member of that tribe pointing to something that the anthropologist immediately identifies as a running deer. The anthropologist is fairly safe in assuming that the informant also thought of pointing to a running deer (and not a moving piece of deer, a shifting pattern of deer fur and grass, etc.). This is so even if the informant also believes that the deer is some other person’s ancestor. If this were not the case—if radical indeterminacy were an omnipresent possibility—then anthropology would be impossible and ethnography could be only fiction.
Before learning to talk, children first learn to follow gaze, engage in joint attention, contingently interact with others to achieve goals, and so forth.
Evolution and Cognition ❘ 48 ❘ 2001, Vol. 7, No. 1
Those who argue that language is acquired through social interaction might be tempted to claim that these facts undermine the modular model of language (SHANKER 2001). Yet, most of the work in this area—called the child’s ‘theory of mind’ or ‘folkpsychology’–focuses on many of the same sorts of questions and approaches that generative grammarians do: How is the child able to reliably infer such rich mental structures about other minds from a few gestures and without mastery of language? And how is it that children also infer that people’s mental structures (intentions) can cause others to act a distance (without any physical contact)? The emerging consensus in the field is that children are biologically endowed with a ‘theory of mind’ (ToM) that matures in predictable ways over the first three years of life (LESLIE 1994; BARON-COHEN 1995; SPELKE et al 1995). Some features of developing ToM interface with maturing features of developing LS in systematic ways that are only now being experimentally studied. This is one of the most exciting and productive research areas in cognitive and developmental psychology (to judge from journal publications and grant funding), and also one of the most thoroughly ‘modular’.
Interactions between modular cognitive systems are complex and difficult to tease apart, perhaps more so than interactions among various bodily systems. For example, belief in supernatural agents, which characterizes all religions in all societies, involves a host of modular expectations and interfacing: folkmechanics, folkpsychology, folkbiology, primary and secondary emotions, predator–prey schema, and so forth (BOYER 1994; BARRETT/NYHOFF 2001; ATRAN in press). Nevertheless, predictive theories are being proposed and empirically tested.
Are Modules Adaptations or Evolutionary By-Products of Adaptations?
Leading evolutionary biologists, such as Stephen GOULD and Richard LEWONTIN (1979) describe the products of higher-order human cognition as evolutionary ‘by-products’ or ‘spandrels’. This is a stance P&P adopt. A spandrel is an architectural term for the structural form or space that arises as a necessary concomitant to another decision in design, and is not designed to have any direct utility in itself. For example, the space beneath a flight of stairs is a byproduct of constructing an inclined stairway rather than a vertical ladder. The fact that people might subsequently use this ‘leftover’ space for storage does not entail that the space was designed to be a storage space.
For GOULD and LEWONTIN, higher-order human cognitive structures originated as functionless spandrels that have been subsequently modified under cultural selection rather than natural selection. Biologically functionless, or nearly functionless, spandrels supposedly include: religion, writing, art, science, commerce, war and play. These evolutionary by-products are cultural ‘mountains’ to the biologically ‘adaptive molehill’ (GOULD 1991, pp58–59; WILLIAMS 1992, pp77–79). On this account, evolutionary psychology would have little to reveal about the emergence and structure of such culturally-elaborated spandrels: “The number and complexity of these spandrels should increase with the intricacy of the organism under consideration. In some region within a spectrum of rising complexity, the number and importance of useable and significant spandrels will probably exceed the evolutionary import of the primary adaptation” (GOULD 1997, pp10754–10755; cf. FODOR 1998).
GOULD’s account of the emergence of distinctly human cognitions as spandrels of a big brain is hardly convincing. The very notion of a ‘big’ or ‘large’ or ‘complex’ brain is too vague to empirically constrain the evolutionary story about how it might have evolved. The big brain is taken as an adaptation from which all cognitive spandrels arise. But an adaptation to what? A design for what? What are its evolutionary-relevant computational structures? Big brain stories tend to be even broader in scope and handwaving than language–evolution stories. Human brains supposedly broke away from ape brains under selection pressures that run the gamut from runaway social competition (ALEXANDER 1989), to gut reduction (AIELLO/WHEELER 1995), to hunting large game (HILL 1999), to niche construction (LA-LAND et al. 2000), to runaway sexual selection (MILLER 2000), and so on. Alternatively, the big brain primarily evolved as a conduit for culture (HARRIS 1975), as a vehicle for language (JERISON 1976), or as a host for the independently evolving ideas, or ‘memes,’ that compete to colonize it (DENNETT 1995; BLACKMORE 1999). Finally, the big brain may have evolved under any number ‘positive feedback’ pathways involving some or all of the factors mentioned.
But even if true, and even if we knew the reasons (selection pressures responsible for) why, we are unlikely to learn anything of particular interest from all this about how the mind works. The notion of a big brain is as uninformative about cognitive structures and functions cognition as the notion of big body is about bodily structures and functions. Never mind exaptations, such as chins, or spandrels, such as
Evolution and Cognition ❘ 49 ❘ 2001, Vol. 7, No. 1
palm lines. From the fact of a bigger body, what could one possibly deduce about hearts, livers, kidneys, hands, faces, placentas and so on? And never mind exaptations, such as language, or spandrels, such as religion. From the fact of a bigger (or denser, or more folded, or grayer) brain, what could one possibly deduce about perception, emotion, categorization, inference or any of the other capabilities humans share with apes but in more vastly elaborated form? Probably nothing at all.
It may well be true that little insight is to be gained into higher-order human cognitive structures by considering possible evolutionary origins and functions. Adopting the hypothesis of GOULD and LEWONTIN or P&P, which assumes this truth, is practically guaranteed to block insight, whether or not it is true. By contrast, adopting evolutionary psychology’s requirement that candidates for exaptations and spandrels be described, as far as possible, with reference to evolved adaptations, then it might be possible to find out if the hypothesis is true or not. If it is true, then evolutionary psychology would have provided the empirical evidence that shows it to be a significant and surprising scientific insight, and not one that depends entirely on intuition, analogy, eloquence or wishful thinking. If it is not true, then evolutionary psychology will have helped to discover something new about human nature.
Three Examples of Modularity: Naive Mechanics, Theory of Mind, Folkbiology
Ever since CHOMSKY jump-started the ‘cognitive revolution,’ successors to the behaviorists who believed in an all-powerful general thinking device have tried to reconcile CHOMSKY’s insights with faith in flexible intelligence by reluctantly granting some specificity to language, and language alone. But cognitive psychology today concentrates more on discovery and exploration of domain-specific mechanisms than on general-purpose computation. Each such device has a particular ‘content-bias’ in that it targets some particular domain of stimuli in the world (‘set of inputs’): for example, the edges and trajectories of rigid three-dimensional bodies that move by physical contact between them (mechanics), the contingent motion a self-propelled actors that can coordinate interactions without having physical contact (agency), or the behaviors and appearances of nonhuman living creatures (species relations). The particular inferential structure of each domain-specific processor then takes the isolated exemplars (or relatively poor samples) of the stimulus-set actually encountered in a person’s life, and spontaneously projects these relatively fragmentary instances onto richly-structured categories (‘classes of output’) of general relevance to our species: for example, the objects and kinds of folkphysics (naive mechanics), folkpsychology (ToM) and folkbiology. Much work on domainspecificity has developed, and now develops, independently of sociobiology or evolutionary psychology (ATRAN 1989; HIRSCHFELD 1996; KEIL 1989; LESLIE 1994; CAREY/SPELKE 1994; SPERBER 1985).
Within the emerging paradigm of cognitive domain-specificity, there is much speculation and controversy—again, as might be expected in any newly emerging science. For example, there are competing accounts of how human beings acquire basic knowledge of the everyday biological world, including the categorical limits of the biological domain and the causal nature of its fundamental constituents. One influential view of conceptual development in folkbiology has been articulated by Susan CAREY and her collaborators (CAREY 1985; CAREY/SPELKE 1994; JOHNSON/CAREY 1998). On this view, young children’s understanding of living things is embedded in a folkpsychological, rather than folkbiological, explanatory framework. Only by age 7 do children begin to elaborate a specifically biological framework of the living world, and only by age 10 does an autonomous theory of biological causality emerge that is not based on children’s understanding of how humans think and behave.A competing view is that folkbiology and folkpsychology emerge early in childhood as largely independent domains of cognition that are clearly evident by ages 4 or 5, and which may be innately differentiated (ATRAN 1987; KEIL 1989; GELMAN/WELL-MAN 1991; HATANO/INAGAKI 1999).
To address this issue, my colleagues and I carried out a series of cross-cultural experiments (LÓPEZ et al 1997; MEDIN et al. 1997; ATRAN et al. 2001). One set of experiments shows that by the age of 4–5 years (the earliest age tested in this regard) urban American and Yukatek Maya children employ a concept of innate species potential, or underlying essence, as an inferential framework for understanding the affiliation of an organism to a biological species, and for projecting known and unknown biological properties to organisms in the face of uncertainty. Another set of experiments shows that the youngest Maya children do not have an anthropocentric understanding of the biological world. Children do not initially need to reason about nonhuman living kinds by analogy to human kinds. The fact that American children show anthropocentric bias ap-
Evolution and Cognition ❘ 50 ❘ 2001, Vol. 7, No. 1
pears to owe more to a difference in cultural exposure to nonhuman biological kinds than to a basic causal understanding of folkbiology per se. Together, the first two sets of experiments suggest that folkpsychology can’t be the initial source of folkbiology. They also indicate that to master biological science, people must learn to inhibit activation of universal dispositions to view species essentialistically and to see humans as inherently different from other animals (ATRAN 1990, 1998).
A third set of results show that the same taxonomic rank is cognitively preferred for biological induction in two diverse populations: people raised in the Midwestern USA and Itza’ Maya of the Lowland Mesoamerican rainforest (ATRAN et al. 1997; COLEY et al. 1997). This is the generic species—the level of oak and robin. These findings cannot be explained by domain-general models of similarity because such models cannot account for why both cultures prefer species-like groups in making inferences about the biological world, although Americans have relatively little actual knowledge or experience at this level. In fact, general relations of perceptual similarity and expectations derived from experience produce a ‘basic level’ of recognition and recall for many Americans that corresponds to the superordinate life-form level of folkbiological taxonomy—the level of tree and bird (ROSCH et al. 1976). Still, Americans prefer generic species for making inductions about the distribution of biological properties among organisms, and for predicting the nature of the biological world in the face of uncertainty. This supports the idea of the generic-species level as a partitioning of the ontological domains of plant and animal into mutually exclusive essences that are assumed (but not necessarily known) to have unique underlying causal natures. The implication from these experiments is that folkbiology may well represent an evolutionary design: universal taxonomic structures, centered on essence-based generic species, are arguably routine products of our ‘habits of mind’, which may be in part naturally selected to grasp relevant and recurrent ‘habits of the world’.
Evidence for Modularity
The evolutionary argument for a naturally-selected cognitive disposition, such as folkbiology, involves converging evidence from a number of venues: Functional design (analogy), ethology (homology), universality, precocity of acquisition, independence from perceptual experience (poverty of stimulus), selective pathology (cerebral impairment), resistance to inhibition (hyperactivity), and cultural transmission. None of these criteria may be necessary, but the presence of all or some is compelling, if not conclusive. Group selection is not a factor.
1. Functional Design. All organisms must function to procure energy to survive, and they also must procure (genetic) information for recombination and reproduction (ELDREDGE 1986). The first requirement is primarily satisfied by other species, and an indiscriminate use of any individual of the other species (e.g., energy-wise, it does not generally matter which chicken or apple you eat). The second requirement is usually only satisfied by genetic information unique to individual conspecifics (e.g., genetically, it matters who is chosen as a mate and who is considered kin). On the one hand, humans recognize other humans by individuating them with the aid of species-specific triggering algorithms that ‘automatically’ coordinate perceptual cues (e.g., facial recognition schemata, gaze) with conceptual assumptions (e.g., intentions) (BARON-COHEN 1995). On the other hand, people do not spontaneously individuate the members of other species in this way, but as examplars of the (generic) species that identifies them as causally belonging to only one essential kind.
Natural selection basically accounts only for the appearance of complexly well-structured biological traits that are designed to perform important functional tasks of adaptive benefit to organisms. In general, naturally selected adaptations are structures functionally “perfected for any given habit” (DAR-WIN 1883, p140), having “very much the appearance of design by an intelligent designer… on which the wellbeing and very existence of the organism depends” (WALLACE 1901, p138). Plausibly, the universal appreciation of generic species as the causal foundation for the taxonomic arrangement of biodiversity, and for taxonomic inference about the distribution of causally-related properties that underlie biodiversity, is one such functional evolutionary adaptation. But a good story is not enough.1
2. Ethology. One hallmark of adaptation is a phylogenetic history that extends beyond the species in which the adaptation is perfected: for example, ducklings crouching in the presence of hawks, but not other kinds of birds, suggests dedicated mechanisms for something like species recognition. Some nonhuman species can clearly distinguish several different animal or plant species (CERELLA 1979; LORENZ 1966; HERRNSTEIN 1984). Vervet monkeys even have distinct alarm calls for different predator species or
Evolution and Cognition ❘ 51 ❘ 2001, Vol. 7, No. 1
groups of species: snake, leopard and cheetah, hawk eagle, and so forth (HAUSER 2000). Chimpanzees may have rudimentary hierarchical groupings of biological groups within groups (BROWN/BOYSEN in press). To be sure, the world itself is neither chaos nor flux: species are often locally self-structuring entities that are reproductively and ecologically isolated from other species through natural selection. But there is no a priori reason for the mind to always focus on categorizing and relating species qua species, unless doing so served some adaptive function (e.g., it makes little difference which tiger could eat a person or which mango a person could eat). And the adaptive functions of organisms rarely, if ever, evolve or operate in nature as all-purpose mechanisms.
3. Universality. Ever since the pioneering work of BERLIN and his colleagues, evidence from ethnobiology and experimental psychology has been accumulating that all human societies have similar folkbiological structures (BERLIN et al. 1973; BERLIN 1992; ATRAN 1990, 1999). These striking cross-cultural similarities suggest that a small number of organizing principles universally define folkbiological systems. Basic aspects of folkbiological structure (e.g., taxonomic ranking, primacy of generic-species) seem to vary little across cultures as a function of theories or belief systems.
4. Ease of Acquisition. Acquisition studies indicate a precocious emergence of essentialist folkbiological principles in early childhood that are not applied to other domains (GELMAN/WELLMAN 1991; KEIL 1994; HATANO/INAGAKI 1999; ATRAN et al. 2001).
5. Independence from Perceptual Experience. Experiments on inferential processing show that that humans do not make biological inductions primarily on the basis of perceptual experience or any general similarity-based metric, but on the basis of imperceptible causal expectations of a peculiar, essentialist nature (ATRAN et al. 1997; COLEY et al. 1997).
6. Pathology. Cerebral impairments (WILLIAMS syndrome, brain lesions caused by certain types of herpes virus, etc.) suggest selective retention or loss of folkbiological taxonomies or of particular taxonomic ranks. Neuropsychological studies have reported a pathological performance in recognition at the life-form and generic-species levels (e.g., recognizing an item as an animal but not as a bird or robin), and dissociation at the life-form level (e.g.,
not recognizing items as trees). Existing studies, however, do not say anything about the generic-species rank as the preferred level of representation for reasoning, perhaps because of methodology (linked to averaging over items and failure to include sets of generic species) (WARRINGTON/SHALLICE 1984; SAR-TORI/JOB 1988; JOB/SURIAN 1998).
7. Inhibition and Hyperactivity. One characteristic of an evolved cognitive disposition is evident difficulty in inhibiting its operation (HAUSER 2000). Consider beliefs in biological essences. Such beliefs greatly help people explore the world by prodding them to look for regularities and to seek explanations of variation in terms of underlying patterns. This strategy may help bring order to ordinary circumstances, including those relevant to human survival. But in other circumstances, such as wanting to know what is correct or true for the cosmos at large, such intuitively ingrained concepts and beliefs may hinder more than help. For example, the essentialist bias to understand variation in terms of deviance is undoubtedly a hindrance to evolutionary thinking. In some everyday matters, the tendency to essentialize or explain variation in terms of deviation from some essential ideal or norm (e.g., people as mental or biological ‘deviants’) can be an effortlessly ‘natural’ but wrong way to think.
Because intuitive notions come to us so naturally they may be difficult to unlearn and transcend. Even students and philosophers of biology often find it difficult to abandon commonsense notions of species as classes, essences or natural kinds in favor of the concept of species as a logical individual—a genealogical branch whose endpoints are somewhat arbitrarily defined in the phyletic tree and whose status does not differ in principle for that of other smaller (variety) and larger (genus) branches. Similarly, racism—the projection of biological essences onto social groups—seems to be a cognitively facile and culturally universal tendency (HIRSCHFELD 1996). Although science teaches that race is biologically incoherent, racial or ethnic essentialism is as notoriously difficult to suppress as it is easy to incite (GIL-WHITE 2001).
8. Cultural Transmission. Human cultures favor a rapid selection and stable distribution of those ideas that: 1) readily help to solve relevant and recurrent environmental problems, 2) are easily memorized and processed by the human brain, and 3) facilitate the retention and understanding of ideas that are more variable (e.g ., religion) or difficult to learn
Evolution and Cognition ❘ 52 ❘ 2001, Vol. 7, No. 1
(e.g., science) but contingently useful or important. Folkbiological taxonomy readily aids humans everywhere in orienting themselves and surviving in the natural world. Its content tends to be stable within cultures (high interinformant agreement, substantial historical continuity) and its structure isomorphic across cultures (BERLIN et al. 1973; LÓPEZ et al. 1997). Folkbiological taxonomy also serves as a principled basis for transmission and acquisition of more variable and extended forms of cultural knowledge, such as certain forms of religious and scientific belief (ATRAN 1990, 1998).
Consider, for example, the spontaneous emergence of totemism—the correspondence of social groups with generic species—at different times and in different parts of the world. Why, as LÉVI-STRAUSS (1963) aptly noted, are totems so “good to think”? In part, totemism uses representations of generic species to represent groups of people; however, this pervasive metarepresentational inclination arguably owes its recurrence to its ability to ride piggyback on folkbiological taxonomy. Generic species and groups of generic species are inherently wellstructured, attention-arresting, memorable and readily transmissible across minds. As a result, they readily provide effective pegs on which to attach knowledge and behavior of less intrinsically welldetermined social groups. In this way totemic groups can also become memorable, attention-arresting and transmissible across minds. These are the conditions for any idea to become culturally viable (see SPERBER 1996 for a general view of culture along the lines of an ‘epidemiology of representations’). A significant feature of totemism that enhances both memorability and its capacity to grab attention is that it violates the general behavior of biological species: members of a totem, unlike members of a generic species, generally do not interbreed, but only mate with members of other totems in order to create a system of social exchange. Notice that this violation of core knowledge is far from arbitrary. In fact, it is such a pointed violation of human beings’ intuitive ontology that it readily mobilizes most of the
stability derivatively attaches to more variable and difficult–to–learn representational forms, thus enhancing the latter’s prospects for regularity and recurrence in transmission within and across cultures. This includes knowledge that cumulatively enriches (e.g., to produce folk expertise), overrides (e.g., to produce religious belief) or otherwise transcends (e.g., to produce science) the everyday ontology prescribed by our evolved cognitive modules.
Conclusion: Neither Sin nor Salvation, but a Promissory Note
Despite the initial independence of work in domainspecificity, there is now increasing convergence in the ways cognitive anthropologists and psychologists, and evolutionary biologists and psychologists, think about related issues. The general consensus is that domain-specific mechanisms likely evolved over millions of years of biological and cognitive evolution to deal with specific sorts of relevant and recurrent problems in ancestral environments (‘task demands’), such as recognizing inert objects (e.g., rocks), reducing biodiversity to causally manageable proportions (e.g., species), or anticipating agents (e.g., the intentions of potential predators or prey). In the computer lingo that now dominates the field, the ‘computational mind’ consists of a variety of distinct, task-specific information-processing devices. Nothing from work on subcortical emotions or neocortical principles of general association has provided, or hints at providing, the slightest insight into the cognitive structures and processes discovered so far in these emerging fields.
Evolutionary psychology is still in its infancy and is not yet the new scientific paradigm some would love and others hate. Some find the idea of ‘Stone Age Minds for a Space Age World’ bold and irreverent, many find it false and demeaning, others find it ridiculous. As the field stands now, all may garner uncertain support for their position. This new field surely will not solve all of the problems
assumptions people ordinarily make about biology in order to better help build societies around the world (ATRAN/SPERBER 1991).
In brief, modularized structures—such as those which produce folkmechanical, folkpsychological and folkbiological concepts—are special players in cultural evolution. Their native
A Continuing Critique of Evolutionary Psychology
Seven Sins for Seven Sinners, Plus or Minus Two1
Introduction
As the astute reader of our paper hopefully noticed, our overall goal was to help guide evolutionary psychology toward a more productive conceptual and experimental path than has been characteristic of the discipline during its earlier stages of development. The ‘moralistic’ stance we adopted was intended to be an attention-grabbing tactic (as is the current title, which is optimally directed to at least five nonparticipants mentioned in footnote 1, and we could easily add or subtract a few additional sins or sinners, including ourselves). We also chose to be confrontational due to our belief that progress in the Evolutionary Psychology2 project should be
Abstract
Of the many issues raised by commentators, two stand out: A debate about modularity and the evolution of language. A neocortically focussed developmental systems perspective, coupled with an understanding of the ingrained emotional and motivational systems we share with other mammals, will provide more insights into what types of intrinsic functions evolution actually constructed in the human mind/brain than will any proposed variant of cortical modularity. For instance, it is unlikely that our higher cerebral expansions could develop language abilities without the ancient pre-existing subcortical systems that generate socio-emotional urges and non-verbal, gestural pragmatics. Although humans may well have ‘communicative instincts’ and eventually ‘proclivities towards language acquisition’, there is little evidence they are born with ‘language modules’. It is imperative for evolutionary psychologists to be constrained by the neuroscience evidence garnered from all mammalian species.
Key words
Evolutionary psychology, human nature, sociobiology, affective neuroscience, modulartity, language, group selection, social selection, natural selection.
constrained by the evidential standard common to all of the natural sciences. Perhaps from the perspective of the standard social science model, Evolutionary Psychology has attained a credible paradigm, but this is not the case from a cross-species, neuroethological perspective. There is abundant room for improvement in our quest for a reasonably accurate and integrated view of human mind/brain evolution.
comes to mind/brain issues, no one can yet unambiguously specify the optimal course of investigation, but if obvious mistakes are not corrected at the outset, they will have increasingly deleterious effects as time passes. For instance, the erroneous concept that the whole brain is a general-purpose, associative learning machine, guided simply by reinforcement processes (i.e., the phlogiston of 20th century behavioral psychology), had a conceptually deleterious effect on mainstream psychology. The notion that human cortical evolution has yielded a mind/brain constructed of massive cognitive modules seems to be a mistaken perspective of the opposite extreme (FODOR 2000). A
Unfortunately, when it
more realistic view lies somewhere in between. Both specific and comparatively non-specific processes reside within the human brain, and once they interact during development, especially in higher regions of the brain (e.g., neocortex), the epigenetically molded neural realities cannot be disentangled if one sides with either polar view. All evolutionary psychologists should appreciate that incorporating developmental systems theory as well as data from related species will be absolutely essential for all conclusions—a point acknowledged by several of our commentators (most prominently PITCHFORD).
Most importantly, in the target article we argued that it is counterproductive to generate claims that do not take all the available evidence into account. Non-biologically based accounts for the evolutionary history of the human mind/brain cannot work if assertions are not consistent with existing neuroscientific evidence. Thus, our guiding principle was that it is much easier to steer a corrective course during the early phases of any intellectual journey rather than when the passage is already well on its way. We think most scholars will agree that current revolutionary variants of Evolutionary Psychology, in their enthusiasm to modify the prevailing and highly flawed standard social science model, have also been making a variety of obvious mistakes that now need to be rectified. To the extent that the discipline can achieve corrective action, it will flourish. To the extent that it cannot, it will increasingly become a scientific cult.
One of the greatest risks to future progress is premature closure on conceptual issues that are not yet empirically resolved. It is probably fair to say that would include essentially everything evolutionary psychologists have suggested about the mind/brain (including most prominently the neuro-evolutionary nature of language). This is not as obviously the case in the cognitive, behavioral and affective neurosciences, where the abundance of converging human and cross-species findings gives greater confidence that lasting knowledge bases have been achieved. Evolutionary psychologists should find it extremely beneficial to pay attention to these findings! We were disappointed that there were few comments on the sampler of ‘solutions’ that we shared in the target article, but we were delighted that Russel GARDNER emphasized the importance of such evidence. It will certainly behoove evolutionary psychologists to assimilate the facts that have been accumulated from ‘lower’ species, even though every finding remains open to various alternative neuropsychological and neurobehavioral explanations. Since most individuals in neuroscience are receptive to evolutionary views, it should be informative for evolutionary psychologists to pay close attention to their interpretation of the neuronal evidence. Indeed, this passage is being achieved on some cognitive issues (DUCHAINE/COSMIDES/TOOBY 2001; GALLIS-TEL/GIBBON 2001), but not, to the best of our knowledge, for the abundant knowledge concerning our basic emotional and motivational systems. Such
neglect resembles the disturbing bias evident in the standard social science model, where evidence derived from our fellow species has traditionally been marginalized. Also, since most investigators of human behavior do not have causal manipulative access to the underlying brain systems, they tend to assiduously ignore the relevant neural evidence culled from other species. However, since we are such a contentious and self-centered tribal species, it is hard to coax those interested in more ethereal human issues to pay attention to such fundamentally important infra-human evidence.
Since the publication of our target paper, we have all experienced the impressive publications of the Human Genome Projects (LANDER et al. 2001; VENTER et al. 2001) and many have been humbled by the finding that the best estimate of the total number of genes in the human species was more than halved. Others, like ourselves, have been delighted with the evidence that at a genetic level, we are not all that different from our fellow mammals—a conclusion that has long been obvious to many. Thus, our vast intellectual/ cognitive abilities cannot be explained by a massively more sophisticated genome, but rather by our massively expanded cortex, the mushrooming of which probably emerged through comparatively modest genetic changes rather than any new and detailed genetic constructions. One should repeat this as another mantra whenever one ponders what evolution did or did not construct in the highest cortico–cognitive reaches of the human brain/mind. Although one might fall back on the emergence of new gene–gene interactions and heterochronic neural progressions in the human species, we again submit that the simplest explanation is the flexibility of mind that general-purpose cortical mushrooming provided. The many ancient attentional, emotional and motivational read–only–memory (ROM) type systems we share with other animals now have access to more abundant general-purpose, random–access–type memory (RAM) spaces that allow complex re-symbolizations of our basic urges.
Due to our massive ignorance of how the cortex really works to generate conscious perceptions, memories, plans and intentions, taken in conjunction with the similarities in the local circuits of sixlayered neocortex in all mammals, we believe this RAM-type metaphor should be the default assumption concerning the function of our heteromodal cortices. Although there is no question that cortical areas become specialized for a variety of adaptive and even maladaptive functions during development, to assume that specialized, sociobiology-type modules evolved in the higher cerebral areas via ge-
Evolution and Cognition ❘ 57 ❘ 2001, Vol. 7, No. 1
netic selection (which we believe is essential to entertain any hard form of modularity) is so patently speculative and at odds with hosts of facts that it cannot be deemed a very plausible neuropsychological viewpoint. It is also a societally dangerous perspective, for it does not leave much room for cultural traditions in the molding of very basic behavior patterns (DUGATKIN 2000; DE WAAL 2001). In other words, the biggest danger with Evolutionary Psychology is its failure to acknowledge the massive flexibility and plasticity of the expansive human neocortex, and its concurrent failure to deal seriously with the many functionally dedicated systems (heavily influenced by genetic factors) that do exist subcortically. In saying this, we also recognize the remarkable importance of genetic imprinting in guiding the construction of such brain systems (JOHN/SURANI 1996; KEVERNE et al. 1996).
All too often, extreme evolutionary viewpoints fail to acknowledge our capacity for well-reasoned, well-intended actions within the many complexities of our social worlds. In any event, it should be clear to everyone who has kept up with modern neuroscience that the genetic tethers on our basic attentional, emotional, motivational and memorial processes are as great as those of other mammals, but at the same time our capacity for selfless action is as vast as the epigenetic/cognitive imaginings of our highest cerebral spaces. The essential role of subtle social and cultural processes in human evolution has been massively underestimated by those who merely focus on selfish gene concepts as opposed to individuals and dyads as the true vehicles of evolution.
We will respond to our commentators in two ways. First we will address each individual commentary, with considerable repetition of the most salient points many commentators chose to focus on, so that each section can be read independently. Then we will move on to several key topics that need to be highlighted and re-emphasized in more detail. Again, we undertook this exercise with the goal of achieving the most accurate evolutionary picture of our animal and human nature that is currently possible, and we shall proceed in that vein.
Ullica Segerstråle: A Historical Perspective
We appreciate the excellent framing for this debate that SEGERSTRÅLE provided. Her comments help put the controversy in an appropriate historical context. Her superb recent coverage of the sociobiology wars (SEGERSTRÅLE, 2000) has nicely set the stage for a better appreciation of the present skirmishes. We wholeheartedly agree with most of the themes she shares, but would quibble with some of the particulars. For instance, we are not adverse to attempts to “find out about the functioning of the mind/brain by asking questions about adaptations and design”. Our objection is only if such questions are asked without considering the understanding that has already been attained, as it has in abundance with regard to our basic attentional, emotional, motivational and memorial abilities. Moreover, we are happy that SEGERSTRÅLE recognizes the need to incorporate ethology into the foundation of psychological thinking. We disagree that ‘real science’ is done at the ‘anatomical, physical level’ and greatly admire theoretical perspectives. Indeed, technologies such as fMRI and PET imaging only yield correlations, not causal relationships. Causation can only be assigned within the framework of well-controlled experiments where underlying variables are manipulated by experimenters.. And causal forces are what evolutionary psychologists seek to discover. As SEGERSTRÅLE recognizes in most of her comments, our battle is against a scientific ‘tribalism’ that ignores broad swaths of the relevant evidence that comes from ‘nonverbal communication’ of our fellow animals. She is correct in recognizing that the study of such shared behaviors, especially their neural substrates, will provide the needed bridge between the discussion of such issues in humans and the detailed analysis of the underlying neural causes in other animals.
The case of human facial and bodily expressions is pregnant with historical antecedents (DARWIN 1998) and we simply note that such processes obviously have dedicated neural circuits that evolution provided as tools for negotiating social interactions. The nature of these systems, especially in terms of overall bodily controls, is not all that different in humans and other animals (although we typically have much richer facial and vocal expressions). However, while most other animals only have an instinctual route to such expressions, we humans also have learned volitional, cortico–cognitive routes that are not directly associated with our genetic heritage. Thus, even when damage to those higher areas leads to an incapacity in the ability ‘to choose’ emotional expressions, the arousal of subcortical emotions still generates overtly expressive emotional tendencies on both face and body (RINN 1984; BOROD 2000). Cortically-mediated facial expressions are probably not controlled by geneticallydictated ‘modules’, although there may be intrinsic,
Evolution and Cognition ❘ 58 ❘ 2001, Vol. 7, No. 1
perceptual/motor proclivities enabling the cortex to mirror perceived actions (GALLESE/GOLDMAN 1998).
SEGERSTRÅLE correctly points out that we do “accuse the evolutionary psychologists of ignoring a whole body of evidence that actually exists”. SEGER-STRÅLE also suggests possible reasons for why evolutionary psychologists have failed to include data from comparative neuroscience in their recent syntheses (COSMIDES/TOOBY 2000). While we agree that nurturing a new scientific field is taxing, we feel that failure to include the large body of neuroscientific data in attempting to explain the ultimate biological causation of human behavior cannot be attributed to the business of a ‘multi-front battle’—it is simply neglect (probably arising largely from the traditional social science bias which tends to intentionally ignore facts derived from other animals). We also appreciate that in doing this, evolutionary psychologists are no different than many other cognitive psychologists who developed new and dubious anthropocentric, intellectual traditions after they became fed up with the all too many abundant sins of many 20th century behaviorists (for an overview, see PANKSEPP 1990).
We sympathize with the multi-front battle that Evolutionary Psychology presently has to wage with many other disciplines. We who believe that studying animal mind/ brains will tell us a great deal about human nature have an even larger challenge to overcome. Our battle is against the prevailing anthropocentric perspective of evolutionary psychology, not to mention psychology in general. Once it is accepted that many of the sociobiology-type systems that humans possess reside in homologous subcortical areas we share with other animals, the temptation to find ‘modules’ in the higher reaches of the human mind/brain should diminish. This is the neurodevelopmental issue (and blessing) that liberates us from any rigid determinism. Again, we have no doubt that there is a great deal of sensory and motor specialization in the cortex, including face-, odor-, posture-, touch-, and voice-recognition dispositions, but we see no evidence that these functions are sociobiological in any deep sense of the word, nor are they uniquely human. Yes, language comes closest to a cortical, sociobiological ‘module’, but let us appraise the evidence cautiously. As SEGERSTRÅLE (2000) understands as well as anyone, much of the confusion in this area has arisen from our political urges and convictions (often unstated) rather than from our willingness to confront all the evidence in unbiased ways. As she says, it is indeed time to “relax the modularity idea”.
Linda Mealey: What’s in a Word?
Linda MEALEY provides a useful and balanced overview of the general arguments, as well as various particulars. She gracefully navigates the pragmatic middle course, highlighting important empirical and conceptual landmarks along the way. We will only reinforce her views by elaborating on a couple of related topics that concern us most. We also have considerable concern with her advocacy of the traditional ‘modular’ point of view that, in our estimation, adds nothing substantive to an evolutionary analysis of brain functions (even though it may have seemed like a good semantic way to start in the midst of massive ignorance).
Of all the ‘sins’, the species-centric perspective is foremost on our minds. It is simply unconscionable, considering all that we presently know about the genetic and epigenetic construction of the brain, that throughout the past three decades practically all variants of psychology have ignored the evidence gained from our fellow creatures (at least as a function of published words). The failure of academic psychology to assimilate information from the neuroscience revolution has been documented several times via citation analyses, and many of us believe that psychology simply cannot be a solid basic science unless it begins to incorporate such issues into its framework.
This complaint should be especially poignant for evolutionary psychology, for many of our mind/ brain abilities are shared with other animals. Even though we clearly outdistance most other species in the complexity and refinement of our cognitive abilities, To the best of our knowledge, the mushrooming of the requisite human neocortex occurred in response to fairly minimal genomic changes. In other words, our cortical expansions are the result of a relatively modest number of heterochronic genetic/regulatory changes rather than any type of major re-engineering that accommodated new and distinct cerebral skills (even though such skills most assuredly do emerge from the epigenetic interactions of our massive brains with our bodily abilities). In short, the highly specialized functions of the oldmammalian brain, situated largely subcortically, took most of evolutionary time to be constructed. If we truly want to scientifically understand the foundation of human nature, a proportionate amount of research effort should be devoted, in psychology, to elucidating these functions. Instead, within psychology such efforts have been a drop in the bucket. Only a few investigators gathered around the deep
Evolution and Cognition ❘ 59 ❘ 2001, Vol. 7, No. 1
well of psychological ignorance (see FODOR 2000) are paying attention to how rapidly other relevant neuroscientific wells are filling with substantive knowledge for understanding the human condition.
All of this important mind/brain work has been conducted without the use of terms such as modularity. Although MEALEY is correct that our view does accept the importance of brain systems, networks and circuits as being of foundational importance for understanding mind, for many of us, the term modularity is a pretentious new-comer that adds nothing to any ongoing analysis. Indeed, it gives us faulty images of ‘encapsulated’ information processing systems that greatly oversimplify highly interactive mind/brain functions that are most rightfully analyzed in neurobiological terms. A similar mistake was made in early neuroscience when ‘centers’ were used as common concepts, but that was soon dropped (except among simplifiers and popularizers) when it was realized that all psychological processes emerged from distributed circuits that interacted with many other functional systems. Since evolutionary psychologists need to come to terms with the organic processes of the brain, they would be wise to discard terms that serve no useful empirical or didactic purposes.
Even though we should certainly not ‘speak of nature versus nurture’ or ‘emotion versus cognition’, we also should not forget that such distinctions serve as heuristics for certain research endeavors, especially those that dip below the surface of psychological experience, to the neurobiological substrates from which such processes arise. Although genes obviously cannot act independently of environments, genes can guide the creation of an enormous variety of creatures and a diversity of mind/brains within a wide range of supportive environments. The distinction between emotion and cognition helps those of us working at the level of the brain to focus on the nature of affective brain processes that desperately need more empirical attention (PANKSEPP 1990, 2000a, 2001a). However, if we only study the psychological/behavioral responses of organisms, then such distinctions seem difficult to justify and investigators may prematurely discard them, although they are still useful in other (e.g., neuro-evolutionary) contexts.
MEALEY eloquently argues that the worth of our psychological concepts ultimately hinges “not on their ultimate ability to ‘reflect’ reality, but their practical value”. She asserts that “if their use encourages inquiry, they are a good thing; if they discourage inquiry, they are a bad thing”. Being pragmatists, we agree with that, but would demure a bit when we consider the neurobiological grounding that is absolutely essential, each and every moment, for our status as creatures of the world. Psychologists are not accustomed to recognizing that the neural analysis provides a vast, new world of ‘reality’ testing. Once we begin to consider all aspects of the systems in which we are interested, we will need many new concepts and perspectives to parse ‘reality’ and we will have to discard others. In neuroscience, the concept of modularity is a non-starter, for it provides no new and useful distinctions. At best, it seeks to replace more informative terms that have been used for decades (e.g., circuits, systems, faculties), while bringing along a great deal of extra and unneeded conceptual baggage. On the other hand, concepts that have long been discarded, such as ‘drives’ may need to be resurrected, for it seems clear from the neuroscience evidence that as we manipulate basic motivations, synthesis of various neuropeptides is genetically induced in the brain to facilitate the energization of specific behavioral tendencies (PANKSEPP 1993, 1998a). We certainly support pluralism in psychological research, but not of the variety that neglects (and too often shuns) the main organ of mind and behavior.
Carlos Stegmann: A Philosophical Perspective
From a philosophical/humanistic perspective we sympathize with the views advanced by STEGMANN. From a practical scientific point of view, we do not, for such holistic views often frustrate any systematic experimental analyses of brain functions. A satisfying holism that postulates different levels within hierarchical systems reflect ‘one–and–the–same thing’ does not advance the analytical discriminations that are essential for the scientific enterprise. Also, we note that there is massive aversion within the psychological community for accepting the idea that within the historically layered and blended complexities of the evolved nature of the mind/ brain “each stratum must be reducible to the laws which apply in the hierarchically lower strata”. Although we do believe there are bound to be strong relations among such strata within the brain, we prefer to see them through the modern philosophical lens of supervenience—whereby lawful relationships at one hierarchical level will be systematically linked, but not completely reduced, to lawful relationships at lower levels (KIM 1993).
Evolution and Cognition ❘ 60 ❘ 2001, Vol. 7, No. 1
We also acknowledge STEGMANN’s concerns with our advocacy of ‘group selection’, for we do believe that pro-social processes must ultimately be instantiated at an individual (and perhaps dyadic) selection level. We raised this issue partly as an enticement to radical selfish-gene selectionists, assuming they are still out there, rather than from any deep conviction that group selection, as traditionally understood, is a robust vector in animal evolution (see “Group Selection” section below)
Our advocacy is based on the recognition that in the evolution of social processes, brain mechanisms may have emerged which only operate optimally within certain social contexts and some may be sufficiently general that they may provide a competitive edge even in the context of outright altruistic behaviors. For instance, since our social bonding mechanisms appear to be designed in such a way as to be based more on learning than intrinsic kin-recognition, it is easy to envision why humans may adopt the children of strangers, even though that is not to their direct genetic advantage. It may simply be to their emotional advantage, even though it may be easy to theoretically (and in our estimation arbitrarily) postulate that it may provide some potential genetic advantage because of the massively shared genetic heritage that all humans share. In any event, we believe that affective feelings are the main psychological currency which guides the behavior of individuals in genetically advantageous (i.e., survival and reproduction promoting) directions.
We look forward to reading STEGMANN’s more comprehensive description of his position, and we will be especially interested in seeing how his ideas might be subjected to empirical tests. Without clear predictions, all views, no matter how holistically attractive they may seem at semantic levels, will remain scientifically sterile. We trust that STEGMANN has taken his interesting analysis to the predictive level. Explanation and synthesis without the precision of new predictions can all too easily appeal to our intellectual–aesthetic senses while not contributing much to the rigor of our analytical endeavors. Indeed, that is one of the ‘siren-song’ appeals of Evolutionary Psychology—it provides such a simple and outwardly credible conceptual umbrella, that often the reductionistic strategies we must develop to scientifically understand the true nature of the underlying processes become of secondary importance. The goal of our article was largely to bring various neglected scientific issues to the foreground of discussion.
Russel Gardner: A Neuropsychiatric Perspective
In his commentary, GARDNER highlights the desperate need to fully integrate perspectives for the study of matters concerning the brain and mind. Typically they are still considered separately, and one of the major challenges he poses is to build bridges that link those disparate realms in ways that are subject to scientific evaluation. Of course, that has been the main goal of affective neuroscience (PANKSEPP 1998a). Indeed, the cardinal scientific motivation of the senior author has been to develop credible scientific structures that may help highlight the deep nature of psychiatric/emotional disorders in the human species. This project, at its outset, recognized that there seems to be no other robust scientific path to understanding the underlying emotional and motivational dynamics of the human brain than through the detailed analysis of homologous processes in related animals. The details of the human brain are simply too inaccessible, but faith in the existence of deep evolutionary homologies has already paid off handsomely in biological psychiatry (CHARNEY/NESTLER/BUNNEY 1999), and it will continue to do so at an ever increasing pace as investigators learn how to effectively modulate neuropeptide systems (PANKSEPP 1993, 1998).
The supposition that we can understand the essential outlines of our own evolved emotional processes by studying the brains of other animals has not traditionally been well received by the psychological community. For many, it remains too much of a theoretical leap to believe that other animals experience emotions. However, on the basis of evolutionary considerations as well as massive anatomical and neurochemical homologies, this possibility is now a solid hypothesis. It is also one that can now be experimentally tested because the neurochemical discoveries in other animals can lead to pharmacological interventions that generate predictions and can be evaluated in humans. For instance, the abundant work on corticotrophin-releasing factor implicating it in anxiety disorders and depression, has led to concordant findings in humans (NESTLER 1998). There are many other examples (CHARNEY et al. 1999).
GARDNER also provides a didactic service by highlighting the history of such approaches in the psychiatric community, noting how ‘selective allegiances’ among powerful academic circles have probably retarded the bridging work that needs to be done. The emergence of evolutionary psychiatry, partly under the umbrella of the Across-Species Comparisons and
Evolution and Cognition ❘ 61 ❘ 2001, Vol. 7, No. 1
Psychopathology (ASCAP) group, has been a clear force in providing new perspectives for future developments in biological psychiatry. While DSM-IV diagnostics have stabilized into a practical and economically useful way to deal with mental disorders in our society, it should not be seen as anything close to the final scientific word. The goal now should be to link many emotional disorders to the activity of specific brain systems. The conceptualization of the basic emotional systems of the brain (e.g., PANKSEPP 1998a, 2000b, 2001b) should be an essential component of future understanding in this area.
GARDNER also highlights several higher-order emotional issues that should be fascinating to evolutionary psychologists. The ‘alpha communication’ style that is evident in manic patients is also evident in many scientific discussions. Given the choice, practically all scientists and scholars are more eager to talk about their own work and ideas than to listen to the stories of others. This urge to talk rather than listen to the next fellow is a very prominent feature of our species and it needs to be explained by the evolved features of our communicative apparatus. We suggest that such urges are much more dependent on subcortical emotional systems (e.g., dominance mediating ones) than intrinsic ‘drives’ of cortico–cognitive brain systems that epigenetically specialize in language skills. We note that modern neuroscience has demonstrated, in abundance, the extent to which seemingly pre-ordained cortical functions can be taken over by other functions, as MEISENBERG asserts, in a ‘guns for hire’ fashion.
In any event, there will be many interesting clinical and evolutionary psychological predictions to be made once we fully understand the primitive sources of our emotionality. It is noteworthy that groups such as ASCAP have emerged to provide “understandable, unfanciful, common-sensical and data-based explanations” of human behavior. In this vein, we also note that the recent emergence of a neuro-psychoanalytic movement that has deep respect for the primitive emotional systems that all mammals share, is another harbinger of a new age of reason in the way we study the human mind/brain (see http://www.neuropsa.com). It also is not shy about fully considering evidence collected from our fellow species.
Gerhard Meisenberg: Modularity is Alive and Well… Supposedly
MEISENBERG seems to agree with our overall thesis and takes the opportunity to provides a wide-ranging analysis of instances of cortical functions that need more attention than we provided, as well as questioning some of our specific conclusions. This represents the type of discussion that needs to be expanded in evolutionary psychology and the key issue from our perspective, once again, is the degree to which higher neocortical functions are modular (genetically-dictated functions) as opposed to experientially modularized (epigenetically derived specializations).
We have no doubt that a great deal of the neocortex becomes highly specialized (‘modularized’?) as a function of experience, explaining the power of the Kennard Principle in neurology (functional recovery is more complete following brain damage in youth than in old age), but aside from a variety of discrete motor and sensory/perceptual processes (many of which MEISENBERG elaborates upon, with which we have no disagreement), we still want to suggest that no sociobiology-type, cognitive ‘modules’ are yet close to being demonstrated (as our original intent was to distinguish examples such as facial/emotional recognition and appreciation capacities from the more cognitive types, which may not have been sufficiently clear in our target article). Certainly, very basic sensory and motor processes became specialized in the neocortex of ancestral mammals a very long time ago, but our main point was that there is essentially no evidence yet that modularized, sociobiological cognitive adaptations have emerged in the neocortex since our neocortical divergence from ‘chimp-brained’ anthropoids some two million years ago. It presently does not appear that most of what evolved in the neocortex since then was intrinsically modular, but rather, general purpose tissue that could become modularized under the influence of environmental factors interacting with functionally dedicated limbic and more primitive subcortical network functions as organisms mature. To our knowledge, there is no credible evidence against this type of epigenetic–developmentalist view. It is of course important from a neuropsychological perspective to analyze these developmentally emergent specializations since they are incredibly important in neurological practice. However, in our estimation, such evidence does not really relate directly to the issue of evolutionary modularity.
We have no conceptual disagreement with MEISENBERG when he discusses the emergence of smiling (which can actually be exhibited by newborn infants), but we disagree with the suggestion that the relevant neurology is merely concentrated in the cingulate cortex. There is as much evidence that the insula and several other cortices participate
Evolution and Cognition ❘ 62 ❘ 2001, Vol. 7, No. 1
in emotional facial expressions (BOROD 2000; LANE/ NADEL 2000) and we are tempted to look for the executive influences over facial emotional displays even in lower areas of the brain, as seems to have already been demonstrated for taste induced expression in a variety of animals (BERRIDGE 2000; STEINER et al. 2001).
We agree that a strict dichotomy between cortical and subcortical processes is ‘too simple’ and was used as a convenient didactic device for efficient communication. However, we believe the overall distinction, with some obvious caveats, remains both conceptually and experimentally ‘useful’. Again, the Kennard Principle tends to be less manifest following damage to many subcortical areas where early damage is generally more devastating than damage occurring later. This is probably because the essential subcortical functions organisms are born with have already been partially remapped to higher areas as a function of hierarchical neural development. Clearly, there is a continuity of functions along the whole hierarchical gradient of the neuroaxis, going from specific hard-wired functions to ever increasingly flexible soft-wired (developmentally programmed) ones, but the simplifying dichotomy forces us to face up to the important fact that there are an enormous number of genetically preordained functions in subcortical regions and a comparative paucity (at least in terms of discrete functions per unit of brain matter) in neocortical ones.
Our main complaint is that the semantics of modularity are very misleading, especially when such entities are seen as ‘encapsulated’ special purpose, cerebral functions interacting in linear cascades. There are no such modules, only highly interactive circuits and widely distributed networks with many overlapping functions (i.e., the semantics of modularity adds nothing to our capacity to discuss brain functions that is not already available in more classic neural systems terminology). These networks are obviously massively interactive in divergent, convergent and reentrant ways, with feed-forward and feed-back components that make any linear processing image essentially wrong and not even heuristic (FREEMAN 1999). At the same time, in the midst of such complexity, we strongly feel that productive investigators will have to continue to invest in simplification, generally recognizing the strategic power of Descartes’ 3rd rule of science—to think in an orderly fashion when concerned with the search for truth, beginning with the things which were simplest and easiest to understand, and gradually and by degrees reaching toward more complex knowledge, even treating, as though ordered, materials which were not necessarily so. We think most investigators realize that the scientific enterprise would be immobilized if there were no attempt to simplify the true complexity of nature.
Perhaps our strongest disagreement with MEISEN-BERG is his contention that the evidence at the subcortical level of analysis is not as robust as we make it out to be. In our estimation, the evidence for subcortical systems (not ‘modules’!) is enormous (PANK-SEPP 1998). Of course, there is room for controversy about the interpretation of specific findings, so let us simply respond to a few raised by MEISENBERG: (1) Although there are abundant debates about details, our current understanding of the mesolimbic appetitive motivational systems (not ‘pleasure’ systems) is truly remarkable, and the role of negative affective processes in engaging such systems have been dealt with in theoretically coherent ways (for overview, see IKEMOTO/PANKSEPP 1999). (2) In mammals, serotonin is a general state control system that modifies every behavior ever studied and it is possible to envision that this system primarily helps keep information channels of the brain reasonably well segregated from each other and helps tone down the impact of all emotionally challenging stimuli (PANK-SEPP 1982, 1986). (3) In addition to the fine work on fear that MEISENBERG cites, there is abundant evidence for subcortical anger–rage processes, sexual systems, nurturance systems, as well as those for separation– distress and play (albeit all these urges are effected through the emergence of cortico–cognitive strategies). (4) The intensity and time-courses of social bonding does vary as highlighted by MEISENBERG’s mention of the now classic differences between the solitary montane- and the highly social prairie-voles, but so far these seem to reflect differences in the underlying intensities of socio–emotional systems rather than difference in kind. (5) There is also abundant evidence for very specific systems to regulate energy balance, water balance, ionic and thermal balances, not to mention various hormones and gases. All of the above issues are extensively discussed in PANKSEPP (1998a).
In sum, we think MEISENBERG’s arguments add important dimensions that need to be considered in the maturation of evolutionary views. We agree wholeheartedly with his striking image of many higher areas of the brain being ‘guns for hire’ which is a striking metaphor for general-purpose learning tissues. Again, we disagree with his assessment that the evidence for subcortical systems remains tenuous, but admittedly, the ongoing research efforts on those issues remain too modest and an enormous
Evolution and Cognition ❘ 63 ❘ 2001, Vol. 7, No. 1
number of details remain to be empirically resolved. However, the evidence that these systems are decisive for the sociobiological competence of animals is rather overwhelming. May we simply recall a blatant early example—a description of cats that had received complete lesions of the periaqueductal gray matter of the midbrain: “If the lesion is extensive, the cats lie inert, silent and flaccid as a wet rag. They never again show any interest in food, but must be nourished by stomach tube. They have no spontaneous activity. They will sometimes swallow if milk is allowed to run down in the back of the throat. Irritation which would cause immediate outcry from a normal cat provokes only feeble movements of the head or limbs.” However, after a few days of recovery, an animal “begins to right itself, first the head and then the entire body. Body reactions are elicited normally. It may even walk slowly about on occasion if stimulated, but has no spontaneous activity, attends to nothing in its environment and never feeds itself” (BAILY/DAVIS 1943, p306). Nothing comparable can be obtained with even extensive damage to the neocortex.
The intrinsic ‘vitality’ of the subcortical terrain, including its importance for the direct generation of a large variety of affective states, should not be underestimated if we truly want to understand basic psychological processes. The capacity of the cortex to ‘learn’ new strategies based upon the dictates of the primary-process emotional and motivational mechanisms of subcortial areas is also truly remarkable. However, any claim that an abundance of Swiss pocket-knife type, intrinsic, inclusive fitness types of cognitive strategies have evolved within the circuitry of the neocortex during the last few million years of human brain/mind evolution remains genetically improbable and certainly unsubstantiated.
We welcome anyone providing evidence for such possibilities, but in their absences, we need not be convinced of an intrinsically modular organization simply through the recitation of well-established neuropsychological specializations that are the stock in trade of neurologists working on the mature human brain. We think all neuroscientists accept that the mature human brain has abundant specializations, as does, to a much lesser extent, the newborn infant, but we may ultimately find that many of these ‘genetically provided tools’ are, in fact, developmentally dependent on the specializations that exist subcortically in infants. Of course, all intrinsic faculties of the mind/brain get massively refined by the experiences of each and every child. The plasticity clearly forces us to consider that the nature
of our socio–cultural environments that we construct for our children are more important than speculations about that stratetgies that EEAs may have programmed into those neuronal spaces (PANKSEPP 2001a). Thus, an evolutionary psychological analysis has yet to add anything to the well-established neuropsychological evidence and it now seems, all too often, to be diminishing our appreciation of the vast plasticity of human potential.
Ian Pitchford: A Challenge From an Insider
Although we acknowledged the probable existence of a variety of specialized functions in the neocortex (even though, once again, the term ‘module’ does not help us think about them in any scientifically useful way), we did strongly argue for the importance of several subcortical systems that mediate emotions and motivations, and allow the cortex to function properly. To the extent that GARDNER sees hope in the neuroevolutionary psychobiology program of affective neuroscience, PITCHFORD seems to dismiss it as a misguided enterprise. However, his remarks were restricted to a small subset of issues that we addressed. Moreover, being the guiding light of an evolutionary psychology list server (nibbs-newsletter@yahoogroups.com associated with http://www.human-nature.com/darwin/index.html), PITCHFORD may have been tempted to give us a stronger tongue-flogging than he otherwise might have (at least that is suggested by some personal communications: vide infra). Accordingly, we see his comments partly as a ‘tribally’ motivated attempt to capitalize on select aspects of the present debate by setting up unrealistic polarities while ignoring many of our main arguments. In any event, he discussed only one of our seven criticisms, namely the “search for adaptive modules where there is only general-purpose computational space”, using language processing and the role of developmental programs as his main exemplars.
For didactic purposes we constructed perhaps too strong a dichotomy between subcortical and cortical functions (as already indicated in our response to MEISENBERG), but that is easy to do when one has personally studied how behaviorally robust and outwardly normal certain animals are after being surgically deprived of their neocortical tissue soon after birth (PANKSEPP et al. 1994). Comparable damage to subcortical areas yields a terminally debilitated animal (e.g., see BAILY/DAVIS 1943 description above). In no way is our position ‘dualistic’. The existence of
Evolution and Cognition ❘ 64 ❘ 2001, Vol. 7, No. 1
adaptive, subcortical systems neither precludes or diminishes the importance of cognitive/neocortical processes in guiding animal or human behavior. We postulate that many of the unique human mind/ brain processes arise from dedicated subcortical systems and flexible neocortical systems interacting with each other, as well as human culture. Moreover, we stand behind our conclusion that the epigenetic molding of cortical functions is far greater than the admittedly substantial experiential influences that also affect subcortical functions (for a more developed coverage of our developmental views, see PANK-SEPP 2001a).
PITCHFORD suggests that we have not adequately absorbed the prescriptions of developmental systems theory (e.g., GRIFFITHS 1997; OYAMA 1985)—that we are talking about ‘genetic blueprints’ rather than accepting the full scope of the developmentalist challenge. We only suggest that he re-read what we have actually written about the matter and we suspect he would be hard put to find as strong an advocacy of the brain developmental systems viewpoint as ours among any other evolutionary psychologists. Some of our past writing on the topic can be found in PANK-SEPP/KNUTSON/PRUITT (1998), PANKSEPP/MILLER, (1996) and PANKSEPP (2000a, 2001a). One of our primary theses is that irrespective of the complex interactions between genes and environment, operating systems that give rise to affective states such as anger and hunger (as well as several others) will invariably reside in subcortical areas of human and mammal brains. How these systems are manifested and resolved at the behavioral level will undoubtedly depend on an animal’s life history, but they will invariably be manifest through the emerging plasticity of higher cortico–cognitive brain systems. The subcortical executive systems for emotional/affective processes have persisted because they stochastically support behavioral phenotypes that produce ‘constant, stable outcomes’ as adaptive solutions to environmental and social demands. As PITCHFORD notes, ontogenetic events in the form of gene/environment interactions support a large spectrum of phenotypes. Genes may not be ‘blueprints’ per se, however they can serve as reliable guidelines that promote ‘constant, stable outcomes’ within the general contexts that their corresponding phenotypes were selected.
The blueprint-perspective of genes that PITCHFORD criticizes, especially with regard to its use of the word ‘innate’, is a terribly important issue to clarify. We also do not support the notion that the genetic material contains pre-determined outcomes. However, genes do carry a certain likelihood of contributing to
certain phenotypes. For example, genes that encode serotonin-binding membrane proteins invariably contribute to the expression of natural behaviors like feeding, mating and aggression (LUCKI 1998). The contributions of such proteins are determined by complex gene/environment interactions (LESCH/ MERSCHDORF 2000). In this regard, nucleotide polymorphisms are inherited characters that can appreciably alter a gene’s expression (for instance, see LE-SCH et al. 1996). However, genes are also up- and down-regulated through environment and experience dependent modulation, which can yield similar results (LESCH/MERSCHDORF 2000). The guideline that a genome carries for specifying the amount of serotonin transporter proteins on the membranes of neurons comprising aggression circuitry, for instance, can and will be altered by environmental demands and individual experiences.
As originally stated in the target article, we continue to reiterate PITCHFORD’s affirmation that evolutionary psychology is well advised to root itself in the conceptual framework of developmental systems theory. However, environmentalism is bound to be rather less influential when it comes to the subcortical neural terrain—at least with regard to the types of neural systems that exist, although their intensities will no doubt be influenced by differential early experiences (PANKSEPP 2001a).
As PITCHFORD may have detected, we do disagree with certain variants of developmental perspectives as advocated by some members of the philosophical community, who seem to relegate DNA to less of an ‘informational’ molecule than most biologists are prone to agree. We feel that if one aspires to understand the nature of biologically-evolved functions/ adaptations, then it is wiser to put a majority of their scientific effort into molecular biological approaches and the true study of ontogeny in neural systems rather than strictly to the study of external issues, even though we acknowledge how incredibly difficult they are to extricate.
Moreover, we hopefully all will agree wholeheartedly with regard to the vast plasticity of cortical learning functions. However, as we will note toward the end of this commentary (in the “Behavior-Genetics—An Essential Key” section), carefully harvested heritability data does highlight that the variability of our basic internal dispositions are controlled more by genes than by external environments, while our overt behavior is guided much more by learning. This is an issue that the modularity advocates of evolutionary psychology have yet to effectively discuss or study.
Evolution and Cognition ❘ 65 ❘ 2001, Vol. 7, No. 1
In sum, although we fully subscribe to the importance of developmental landscapes in molding higher mind/brain capacities, we do not agree with the full revolutionary fervor of the ‘Ontogeny of Information’ critique of genetic influences. The genes are more influential in the construction of organisms than the classic OYAMA (1985/2000) type of view seems to accept. We would be surprised if PITCH-FORD would disagree. Surely, we all now agree that genes can do nothing without supportive environments. However, a remarkable amount of organismic competence naturally unfolds from the genome and the resulting internal milieu as long as a minimally supportive external environment is present. There is no comparable robustness that has ever been demonstrated through variations of prenatal environmental factors, at least within a range that most reasonable investigators would deem to be normative. However, in pointing out this obvious fact, we do not think that it forces us into the position of perpetuating the ‘nature versus nurture dichotomy’. As we said in our article, developmental systems theory is an incredibly important concept, but that in no way should be taken to diminish the importance of genetic factors. If PITCHFORD does, in fact, believe that external environmental issues are as important for the functions of subcortical areas as for cortical ones, we disagree with him based on our reading of a massive amount of available neurobehavioral literature. If he believes that environmental factors are more important than genetic ones in molding cortico–cognitive functions, we would agree with him.
PITCHFORD shares an abundance of recent new information about cortical issues, but those do not relate clearly to our claim that much of the specialization of the cortex is due to self-organization through development whereby functionally dedicated subcortical systems interact with a remarkably plastic cerebral mantle. Nothing about the interesting anatomical issues he summarizes relate to that key issue, and in reading his extensive anatomical coverage, we felt he was creating a verbal smoke-screen (or ‘bait and switch’ as Dan DENNET 1995 might say) rather than dealing with the issues we had raised. Those anatomical issues are most interesting and important, but they do not unambiguously relate to either developmental or intrinsic functional issues. Furthermore, we would note that although PITCHFORD cites work that reports humans have a brain size over three times the size of a non-human primate of similar size (RILLING/INSEL 1999), there has been no selective increase of relative frontal lobe size within the human species (SEMENDEFERI/DAMASIO 2000). He does not cite a single functional study that points clearly toward a reasonably self-sufficient, evolutionarilydictated cerebral module that underlies a psychological process that evolutionary psychologists have predicted.
We are also fascinated by the spindle neurons found in the anterior cingulate by NIMCHINSKY and colleagues (1999) and their potential (though yet undemonstrated) role in socio–vocal communicative intent (an anterior cingulate function we noted in our target article). We suggest that of the many specialized cerebral functions that have been discovered by neurologists and neuropsychologically-oriented investigators during the 20th century, none have yet been clarified any further by evolutionary thinking (but see BARKLEY 2001 for some changing contextualizing trends). In any event, it is more important for evolutionary views to generate new neuropsychological findings rather than simply providing a new context for discussing findings discovered by other disciplines. In this realm, the emerging related discipline of neuro-psychoanalysis seems to be farther along in pursuing such issues (SOLMS 1997; SOLMS/NERSESSIAN 1999; KAPLAN-SOLMS/SOLMS 2000) and we believe such contributions highlight the type of work that evolutionary psychologists should begin to pursue in earnest.
As soon as evolutionary psychologists begin to give us new information about the neuroanatomical location, as well as the underlying neuro- and psychodynamics of their evolved ‘modules’, they shall have provided biological evidence for what biological evolution has created in the human mind/brain. In their absence, claims of evolved cortical modularity must be deemed insubstantial, no matter how many individuals have imprinted on the idea. However, we heartily encourage such discoveries and the application of the armamentarium of new scientific tools that are now available for the study of such problems (TOGA/ MAZZIOTTA 2000). But please remember that correlations do not clearly reflect causations.
Although certain human proclivities, such as language and theory of mind abilities, are surely linked to our higher cerebral abilities (GALLESE/GOLDMAN 1998; BARKLEY 2001), it will be equally important to determine how those mind/brain functions are linked to the more ancient systems that we share with the other animals. Indeed, we believe that human facility for spoken language may be deeply linked to evolutionarily-derived, emotional/communicative pragmatics exhibited by other mammals. In our cultural evolution, it is certainly possible that highly-prosodic, sing-song languages emerged long
Evolution and Cognition ❘ 66 ❘ 2001, Vol. 7, No. 1
before we developed the practice of communicating in more measured, less dynamic spoken tones (WAL-LIN/MERKER/BROWN 1999). However, since we know so little about the neurodynamics that create the capacity for speech, we agree with those modern linguists (e.g., LIEBERMAN 2000) who believe that our facility for speech is an outcome of primitive subcortical systems for communicative intent interacting with comparatively more plastic, higher cortical systems through complex developmental trajectories. Yes, human speech functions do tend to get focussed in specific regions of the brain during development, but the question that we must all now consider is how such specialization actually emerges. We think that aside from cortico-limbic participation (e.g., the anterior cingulate systems for communicative intent), most of language may emerge via epigenetic plasticity in fairly general-purpose, but highly multi-modal learning circuits of the neocortex. Certain subcortical systems (e.g., the basal ganglia) may be decisive in the emergence of such higher abilities (LIEBERMAN 2000; BREITENSTEIN, et al. 2001).
We suspect that PITCHFORD would generally agree, even though he is adverse to admitting that subcortical functions are important for evolutionary psychologists to consider for as uniquely a human activity as propositional language. To what extent “language may have adapted to the emerging complexities of the brain rather than the other way around” (p111 of target article) remains an open issue, no matter how strongly certain individuals (including ourselves, of course) believe in certain evolutionary scenarios. And surely PITCHFORD would agree that our ability to read and write, not to mention most of our numerical abilities (beyond primitive counting), are largely culturally induced skills that may have rather little do to with specific evolutionary selection for such abilities (even though we suspect that behavior geneticists could certainly select for such skills by increasing the speed and quality of certain higher cortico–cognitive abilities). If so, are these widespread skills of the members of human societies examples of adaptations, exaptations or spandrels?
To some extent, and understandably, PITCHFORD’s flogging of our view may be based on his failure to be conversant with what the senior author has written in the past (e.g., PANKSEPP 1998a, 2001a). Just to highlight how committed we are to the developmental systems view, we refer PITCHFORD to the senior author’s vigorous advocacy of treating ADHD with environmental approaches, and the ongoing fight against prevailing pharmacological interventions for ‘normal’ childhood impulse-control problems (PANKSEPP 1998b,c). It is quite remarkable to us that in the same sentence, PITCHFORD indicates that the accomplishment of Evolutionary Psychology (i) “compares extremely favorably with that based on the neurochemical individuation of traits and disorders admired by” us, but which have (ii) “produced so many unsatisfactory theories and treatments (VALENSTEIN 1998)”. Is he suggesting that Evolutionary Psychology has also produced an abundance of the latter?
In any event, we believe that VALENSTEIN’s interesting historical analysis of the biological psychiatry revolution was based on an older era of psychopharmacology rather than the future neurochemical revolution we were discussing based on emerging knowledge concerning the highly specific functions of various neuropeptides (PANKSEPP 1993; 1998a). We also suspect that VALENSTEIN’s analysis was also influenced as much by his personal socio–politico–philosophical leanings as his scientific ones (for more on this issue see the “Politics and Tribalism” section). For instance, the senior author’s long-term advocacy of the foraging/SEEKING/expectancy view of lateral hypothalamic reward (PANKSEPP 1981, 1982, 1998a) was never supported by VALENSTEIN, whose work on the plasticity of those systems originally threw this field of inquiry into theoretical chaos (VALENSTEIN/ COX/KAKOLEWSKI 1970). Only when work by his own close colleagues suggested a very similar view (BER-RIDGE/ VALENSTEIN 1991), did this type of concept emerge into more open consideration in behavioral neuroscience (ROBINSON/BERRIDGE 1993). VALENSTEIN has long championed the massive role of learning and plasticity in the emergence of psychological functions in the brain (and has strongly argued against subcortical specificity of functions), and we believe his analysis of the psychopharmacology revolution was motivated to some extent by his philosophical/democratic point of view.
Moreover, we are most certainly not dualists of any stripe and Steven J. GOULD has not been ‘our mentor’ (even though we admire his writing skills and with the help of SEGERSTRÅLE’s (2000) analysis, better appreciate his humanistically motivated willingness to present the other side of the argument in the lucid, energetic and perhaps politically motivated ways that he has). Accepting the vast plasticity of the neocortex does not make us advocates of untenable variants/hangovers of CARTESIAN thought. Our views are thoroughly naturalistic, monistic and materialistic (without abandoning our spiritual/humanistic compass, which we believe is closely linked to our deep animalian capacity to experience pro-
Evolution and Cognition ❘ 67 ❘ 2001, Vol. 7, No. 1
social emotions). We accept that the multilayered complexity of the human mind cannot be understood if we do not come to terms with our ancestral past.
We were dismayed that PITCHFORD purports that we do not subscribe to a hierarchical view of brain organization, intimating instead we advocate a perplexing ‘horizontal layers’ view of brain organization. Obviously, the brain is layered hierarchically, with higher functions critically dependent on various lower functions (please refer to Figures 10.3 — 10.5. of PANKSEPP 1998a), and the flexibility of the lower functions has been increased enormously by the regulatory capacities of the higher functions. In our estimation, it is the modularity view that seeks informationally ‘encapsulated’ evolved units in brain areas that do not have an adequate evolutionary history for their construction, nor an anatomical place or reasonable neurodynamic way to be plugged into the rest of the brain.
A great deal in evolutionary psychology will ride on the exact nature of the massive neocortical mushroomings that characterize the anthropoid/humanoid brain. Whatever specializations exist in the neocortex, there is a good possibility that they are the ones we share with other animals, and the rest is largely a quantitative expansion of these same themes as opposed to major re-engineering of functions. Mere increases in computational space can create seemingly qualitative emergent effects. PITCH-FORD also suggests that “our brains may not simply provide an excess of plastic neurons capable of subserving any function, but may be a solution to the problem of retaining adequate functioning over a prolonged period of time”. This is not an explanation of increasing hominid brain complexity at all. A relatively small, loosely-structured, ungyrified ‘brain’ suits a forty pound lobster of approximately eighty years just fine! ‘Prolonged function over time’ is as creative as any unsubstantiated Panglossian explanation, as it would be an advantageous property of nearly every adaptation. More significant to PITCHFORD’s argument is that we posit different and seemingly unrelated processes for the evolution of subcortical and cortical brain structures. This could not be any further from what we actually think. Natural selection has had free roam in all areas of the brain. However, it should be obvious to all that natural selection has been molding ancient subcortical brain structures for a much longer span of time than those found in neocortical regions of the brain. The more that something has been ‘selected’, the more consistently, efficiently, economically and reliably it will serve its function. We can be quite certain about the genetically ingrained nature of many subcortical functions. We can have no comparable confidence for neocortical ones, especially in humans where early development and cerebral maturation go temporally, hand in hand (DAWSON/FISHER 1994; THATCHER et al. 1996; PANKSEPP 2001a).
Above all, we subscribe to the position that the fertility of ideas is linked inextricably to our ability to predict and explain new phenomena, and we believe that from this perspective, neuroevolutionary psychobiology far outshines Evolutionary Psychology. We are confident that PITCHFORD, were he tallying up the substantive findings and claims, would see the strength of our strategy and also our robust allegiance to a deep evolutionary view of human and animal nature. In any event, while seeking out the weakest chinks in our evolutionary armor, he assiduously avoided the most important aspects of our arguments concerning the core-nature of our emotional and motivational processes. We hope that in the long run, evolutionary psychology will not do the same, even though in the short-run we do anticipate a ‘tit–for–tat’ strategy.
In any event, it should be a ‘no brainer’ for all evolutionists that if we do not understand our ancestrally derived attentional, emotional, motivational and memorial capacities, we can never really understand our own nature. The most dangerous place to seek modularity is among the highly-specialized, neocortical functions of the adult human brain. This may be a bitter pill for most evolutionary psychologists who seem to wish to force a unique human nature onto our species and hence, seem to have an aversion to both animal- and organically-focussed research. Their most robust general principles and phenomena (as well as potential allies) will be found among the subcortical functions of the brain (and presumably among investigators who have studied such functions). They simply need to drop their ‘social science, anti-biological tribalism’ and assimilate the types of lessons that apparently are second nature to their colleagues in ASCAP and human/animal neuroethology societies.
Scott Atran: The Language ‘Module’ Once More
Obviously, all neuoscientifically informed people agree that the human cortex permits levels of abstraction that are not available to any other species. Human language is the most striking example of this ability, as it has attracted more experimental
Evolution and Cognition ❘ 68 ❘ 2001, Vol. 7, No. 1
and theoretical attention than any other faculty of the human mind. Clearly language does not merely arise through SKINNERIAN associations, and we are as attracted to the view of a deep-structure for language as much as anyone, but we agree with CHOM-SKY that it is premature to genetically map it onto neocortical areas of the brain. Of all the non-trivial ‘modules’ that have been proposed (i.e., those beside the straightforward sensory and motor capacities of the brain), those devoted to language are clearly among the most compelling.
ATRAN provides a balanced overview and a vigorous defense of modularity with language as the centerpiece of his argument. We agree with much of his analysis, but we still think that it is wiser to hold off on any strong, genetically-guided modularity view, since the arguments against a uniquely human genetic guidelines for language remain quite compelling. We think many other organisms could also exhibit language if they too had a much more complex neural space for general-purpose, perceptual/cognitive processing within their neocortices and more sophisticated voiceboxes. Yes, we agree that language has become a very adaptive tool for the human species, but so have many of the other emergent properties that culture permits. We think that all may agree that reading and writing are not derived from our genetic heritage, and it is a pity that some of us have caved in so rapidly to the view that our vocal expressions of language, at least in their basic outlines, are constructed in such a manner. We certainly agree that many spandrels can emerge from a vast, predominantly general-purpose learning organ (i.e., the neocortex) which ultimately (i.e., epigenetically) will contain functionally distinct neural circuits.
The most critical issue in our minds is whether a newborn baby has the intrinsic neural circuitry for language or only the intrinsic proclivity to babble while it perceives the world, points to the world, grasps objects in the world, manipulates the world in a self-referential way (all very primitive mammalian abilities) and finally begins to associate these ongoing vocalizations to certain activities in ways that gradually grow into the vast complexities of human language. The only issue that we felt we had raised with this example of modularity was whether our genes contain the rudiments of a uniquely human language, or whether they merely help construct a well-placed voice box and resonance chamber for making complex sounds that can interact with the enormous RAM-space in the cortex, with language as its final product. ATRAN provides a vigorous defense of a sophisticated variant of the accepted
wisdom and we chose, largely for didactic purposes, to take the contrary position that is certainly not falsified by evidence thus far. Our main goal was to highlight the simple fact that in our evolutionary thinking, we should not stray from the available neurodevelopmental evidence to conjectures about the potential evolutionary sources of the final product too quickly. This is a primary reason why we support the comparative study of subcortical processes so ardently, for there is much more assurance of having relevance to our neuro-genetic understanding of human behavior.
We should all be more willing to consider that a great deal of communicative intent and its associated pragmatics may arise from ancient processes we share with other animals. We believe that humans would have no lingusitic competence without the brain systems that exist below the neocortex and if one can appreciate this point, one may also understand why the human neocortex may not require a genetically dictated language ‘module’ to have communicative competence (e.g., BROCA’S and WER-NICKE’s Areas may emerge developmentally because of other compatible multi-modal functions of these tissues). Moreover, we hazard the prediction that once the neocortical circuits for language are completely elucidated, it will be impossible to simulate linguistic competence in a program that instantiates only those higher cerebral processes. In other words, the likely result of this thought experiment would be that by itself, the unique human neocortical architecture could not generate, certainly not motivate, a substantially meaningful human language. Indeed, we note that at least a third of classic KANNER-type autistic children are incapable of speech throughout their lives, although they do not typically have any higher cerebral dysfunctions—it is simply that their higher brain areas are not well connected to subcortical systems (BAUMAN/KEMPER 1995).
Let us be frank about our position: Just as we believe there are many subcortical faculties that are absolutely essential for the types of psychological processes which ATRAN discusses, we also believe that there are bound to be some cortical faculties of considerable importance for the emergence of human language. In describing the cortex as a general-purpose type of ‘information processor’, we are not committed to the view that it is nothing but a general purpose associative tool. Cortical processing is guided by many subcortical processes, and indeed we have previously envisioned that there are cortical ‘spheres of influence’ for higher-order processing of information relevant to various emotional and mo-
Evolution and Cognition ❘ 69 ❘ 2001, Vol. 7, No. 1
tivational survival concerns (PANKSEPP 1988, 1989). We anticipate that those ‘spheres of influence’ may process information in unique ways. We also believe that the frontal lobes have some very general abilities that give it such a unique role in elaborating various executive processes (GALLESE/GOLDMAN 1998; BARKLEY 2001). Indeed, many such executive processes probably participate in so many different cognitive functions that the concept of modularity merely becomes a hinderance in trying to work out how some of the basic cortical processes that help construct so many sophisticated cognitive abilities via the experiences of individuals.
Thus, we are not adverse to studying such higher faculties, but we suggest that evolutionary views have not and are not likely to have any impact on how we empirically study these functions, although they can help loosen up the types of intellectual constipation that have characterized past variants of what might be appropriately called ‘dustbowl cognitivism’ (see BARKLEY 2001 for an example of this laxative experience). In any event, our position was that ultimately, most of the sophistication of the cortical functions is brought to life experientially. An understanding of the neuro-maturational processes that yield well-honed cortical processes will be much, much more, dependent on general brain mechanisms that govern neuronal plasticity than any type of resolved information that is contained in the genes. That was our main point, and we encountered nothing in ATRAN’s well-crafted defense that dissuades us from this position.
We see our disagreement amounting to little more than a matter of emphasis. ATRAN wishes to retain the concept of modularity and sees some type of implicit advantage in that evolutionary view, although he does not suggest how it might be cashed out empirically. We see both of those issues to be superfluous for the pursuit of his faculty-psychology program. We regret that he fails to see the potential power of subcortical, socio–emotional systems in governing many of the processes in which he is interested. Thus, to take his first example of a structural principle, ‘the whole–object constraint’, the understanding of which modularity has presumably facilitated, we point out that the visual system is certainly designed during early development to perceive the objects of the world (this is also evident in other animals). In our estimation, young children attach labels to such objects using social, ‘point and name’ games with caretakers. To underestimate basic social/motivational/emotional processes in such linguistic acquisitions is, we think, shortsighted.
Considering that ATRAN dealt with only one small segment of our critique, we were surprised that he claimed that “there is no empirical evidence whatever to indicate” that our “central thesis is true, and much to suggest that it is false”. If the ‘seven solutions’ we shared are of relevance for human behavior (admittedly most remain to be tested in humans), then a cognitive psychology that continues to neglect such issues must surely be deemed nothing short of myopic. We think that the failure of most commentators to deal with the animal data we provided simply highlights that a great deal of the evolutionary processes that control our behavior appear to be of little interest to evolutionary and cognitive psychologists. We suspect that many of those processes are absolutely essential for the higher processes in which cognitive scientists are so interested and we will only note the disastrous effects that damage to those subcortical systems can have on human mental life (DAMASIO 1999; SCHIFF/PLUM 1999).
As we stated in the target article, while socialization is certainly not a direct evolutionary explanation of human language (as ATRAN also emphasizes), it probably was a selective force for the emergence of communicative intent. We assume that all will agree that relatively little human language would emerge without socialization. We can also agree, that in most people when they have learned language, a great deal of the critical circuitry is concentrated in WERNICKE’s and BROCA’s areas of the left hemisphere, as well as the circuits of the anterior cingulate. Moreover, if all those tissues are destroyed, the child will still learn language by using, presumably, the corresponding systems of the right hemisphere (BATRO 2000 VARGHA-KHADEM et al. 1997). However, the plot now thickens. Children born deaf also aspire to communicate and they do quite successfully, partly by using somewhat different parts of the brain where somatosensory skills are usually elaborated (NEVILLE et al. 1998; PASQUAL-LEONE/TORRES 1993). And where do multiply impaired individuals like Hellen KELLER elaborate their urge to communicate?
We, along with others (LIEBERMAN 2000), believe that so much of language is linked to subcortical and cortico–limbic social urges/pragmatics, that it may be that language emerged from these ingrained systems interacting with our massively sophisticated RAMtype space of the neocortex. The reason the left hemisphere has such primacy in most people is because the left side of the brain is more talented at extroverted social activities (for reasons we will not entertain here), and because WERNICKE’s area is nestled between all of the major perceptual surfaces of the brain
Evolution and Cognition ❘ 70 ❘ 2001, Vol. 7, No. 1
that harvest exteroceptive information (i.e. vision, hearing and touch). The ultimate function of language is to symbolize the world in abstractions and to allow us to develop better internal thought for the pursuit of our many basic goals and externalized strategies for coordinating group activities.
Thus, we are able to buffer information about our role in the world in much more complex ways than any other species, but where is the intrinsic, evolutionarily-selected ‘module’? How could genetic selection have created such dispositions uniquely for the human mind/brain? Where are the genes for language? Might all the pragmatics have already been there before humanoids walked the face of the earth? Might the main thing we needed to cross the syntactic-cultural Rubicon have been a massive endowment of more RAM-type space in the right parts of the brain working within the context of the mounting medium of cultural evolution?
In saying all of this, we must again emphasize that we certainly agree that there are many specialized functions in the adult human cortex. In our original article, we did not aspire to suggest that there were none. Not for a moment do we believe that higher faculties are constructed merely from associative networks. We think that the cortex has many special ways to construct meaning (BARKLEY 2001). There are intrinsic mirroring processes (RIZZOLATTI/FADIGA/ GALLESE/FOGASSI 1996), which may serve as a major vector in the acquisition of language (GALLESE/GOLD-MAN 1998). However, we suggest, along with some others (LIEBERMAN 2000), that the cortex could do none of that—simply none of that—without the support of subcortical systems (and that seems to be a novel perspective for many). We remain perplexed why so many scholars are willing to commit themselves to neurologically unrealistic images of human nature, such as the neurobiological and cortical modularities of evolutionary psychology, which are not yet supported by essentially any sociobiological data. Our main goal was to interject a much greater note of necessary doubt into the intellectual activities of recent variants of Evolutionary Psychology.
We disagree deeply with the ‘separatist’ cortico– cognitivist position advanced by ATRAN. The importance of subcortical and limbic systems in human affairs has been massively underestimated. When he says “nothing from work on subcortical emotions… has provided, or hints at providing, the slightest insight into the cognitive structures and processes discovered so far”, we suggest this view simply reflects a dismissive mode of inquiry preferred by many intellectuals interested in the higher human proclivities and abilities. The subcortical perspective will provide a great deal once individuals begin to study why human minds dwell obsessively on certain themes when they are hungry, thirsty, cold, sexually eager, craving drugs and worrying about their social attachment concerns. There are bound to be higher ‘centers of gravity’ for the cognitive elaboration of such basic functions (PANKSEPP 1988, 1989), perhaps epigenticially magnified or diminished depending on early experiences (PANKSEPP 2001a). The whole field of biological psychiatry has been based largely on manipulating subcortical systems, many of which directly modulate cortical functions. There will be a vast new biological–psychiatry revolution of even greater magnitude when we learn to manipulate other peptide systems in the subcortical reaches of the brain, especially those that control specific emotional and motivational processes (PANKSEPP 1993; 1998a). These molecules will take us to the appetitive core of our existence. We will derive whole new layers of deep understanding about the emotional imbalances that are called psychopathologies. Animal models will help us work out many of the essential neural details of such imbalances, and thereby the sources of our everyday likes and dislikes that continue to be among the most robust sources of personality variability, as well as the basis of so many of our human interactions.
There is an ‘affect logic’ to our cognitive activities that has barely been studied empirically (SHAND 1920; WIMMER/CIOMPI 1995). We intentionally left out one phrase (i.e., ‘or neocortical principles of general association’) in the above quote because we are not as committed to a fully associative mode of cortical processing as some of our commentators wished to impute to us. One can have general-purpose learning functions in the brain which are not simply based on association-network heuristics (FREEMAN 1999). Aside from a few well-analyzed sensory/perceptual and motor processes, our knowledge of how the ‘great intermediate net’ of heteromodal cortical tissue operates remains a great mystery. We are tempted to add that essentially nothing from work of Evolutionary Psychology has provided or hints at providing, lasting insight into the cortico–cognitive structures and processes that truly exist in the human brain. Still, the evolutionary view can certainly help generate such hypotheses (e.g., BARKLEY 2001). If we could drop our destructive species-centrism (which no commentator denied being a problem), we can learn an enormous amount about how the human mind/ brain operates (DE WAAL 2001; GALLISTEL/GIBBON 2001; PANKSEPP 1998a).
Evolution and Cognition ❘ 71 ❘ 2001, Vol. 7, No. 1
Let’s Admit It: We Don’t Know Squat About the Evolution of Language!
We know about three definite things about the evolution of language: (i) The voice box and rest of the upper respiratory apparatus appear to have undergone evolutionary shaping such that we can make a remarkable variety of resonant sounds. Of course, other creatures, songbirds for instance, have achieved similar excellence in sound variety without advancing much toward human-type language or human-type neocortex. (ii) The areas of the left hemisphere that are especially important for language (e.g., planum temporale) are already enlarged at birth and palentological evidences indicates these organs enlarged quite a long time ago—namely, when our brains started to expand beyond the proportions of other great apes some two million years ago (for a summary of that work, see LIEBERMAN 2000). (iii) We can’t recall this one right now, but it does not matter much, for if that is essentially all there is to our knowledge of the evolution of language, aside from our widespread readiness to be awed by this uniquely human skill, the evolutionary evidential base must be deemed slim indeed.
Of course, we are all very impressed that in the absence of brain damage or major sensory deficits (e.g., deafness) language is biased to settle in the left hemisphere in circuits known as BROCA’s and WER-NICKE’S areas. Indeed, people with a college education tend have more robust neurons (i.e., larger dendritic fields) in some of these regions compared to high school drop outs (JACOBS/SCHALL 1993; JACOBS/ SCHALL/SCHEIBEL 1993), and there are many other brain/language correlates (GRABOWSKI/DAMASIO 2000). However, we should hold off on surmising that such language functions reflect biologicallyevolved skills that have any pre-determined and detailed cognitive resolution. After all, these are also among the most impressive pharyngeal-motor and exteroceptive-associative areas, and it seems quite reasonable for language, which allows us to re-symbolize the experience of the individual senses, to find a natural epigenetic home in such areas. Many may be tempted to localize some fine evolutionary ‘needlework’ for the deep structure of language here, but no one is quite sure what one might look for in those locations—anatomically, electrophysiologically or neurochemically—that might yield data consistent with an evolutionary scenario.
Parenthetically, and to reply to ATRAN’s most daring attempt to dispatch out critique (i.e., that there is no evidence for the role of subcortical functions in
cognition), let us simply assert that we have no reason for a comparable agnosticism when it comes to subcortical functions. There is an abundance of anatomical and neurochemical data for a host of functions that speak clearly for strong and powerful homologies across all mammalian species (PANKSEPP 1998a). We can also be certain that these systems have great influences on our cognitive processes: when we have strong emotions and motivations in our minds, related topics are essentially all that we are prone to speak or think about. It is an intellectual tragedy that the cognitive sciences have marginalized these incredibly important aspects of our mental lives. We suspect many loose pieces in cognitive puzzles would fall together well if we would simply focus on the affective sides of human lives a bit more.
But speaking is only one form of communicating and we know that when the most robust neural locations for language processing are damaged, many people are able to communicate quite well again, using different, albeit nearby, regions of the cortex. Sign language seems to have a natural tendency to nestle near the classic left-hemisphere speech areas, but reading is vastly different in deaf individuals (NEVILLE et al. 1998). When people have strokes, language and other functions tend to be restored quite well by unaffected tissues that surround the damaged area by becoming more active during communicatory actions and intents (BUCHNER et al. 1996; NUDO et al. 1996), and language acquisition certainly exercises the brain in most interesting ways (CASTRO-CALDAS et al. 1998). In short, communicative competence can emerge from many areas of the human cortex and all of it requires a variety of subcortical functions about which we know very little (GRABOWSKI/DAMASIO 2000; LIEBERMAN 2000). On the basis of abundant neurological data, there appears to be a major role for subcortical structures in such processes (BASSO/SALA/FARABOLA 1987; CROSSON et al. 1997; BECHTEREVA/ABDULLAEVA 2000). Perhaps this is the third finding we were floundering for earlier, or maybe it was the very important role of prosody (e.g., motherese) in promoting language development in children (FIFER/MOON 1994). But it does not matter, for the evidence supporting evolved cognitive processes of language is tenuous and in the absence of specific genetic details, it is unlikely to get much stronger.
We could carry on further with these lines of thought, but the key point has now been made a few too many times. So let us, ever so briefly, indulge our own evolutionary fancies. The multi-modal association tissue of the cortex that is essential for linguistic
Evolution and Cognition ❘ 72 ❘ 2001, Vol. 7, No. 1
symbolization will work more effectively if there are a greater number of neurons that can ‘crystallize’ their tasks efficiently. Thus, we suspect that the evolutionary advantage may have been for individuals that had more tissue in those brain regions, faster communicating neurons and perhaps the many factors that contribute to an exquisite plasticity that can become readily ‘gelled’. And perhaps little more, except for the fact that when these systems become disconnected from the deeper feelings of the right hemisphere, they tend to confabulate like crazy (PANKSEPP 2000c). It is most remarkable that psychological procedures, such as psychoanalysis, can get behind the confabulatory facade to deeper principles of reality (KAPLAN-SOLMS/SOLMS 2000). If one puts our whole vision of human linguistic nature together, it may help explain why most of us tend to be better speakers than listeners, and why so many of us are eager to speak faster and louder than the next fellow, until there is little more than a blistering cacophony of sounds.… “Till human voices wake us and we drown.” (T. S. ELIOT, The Love Song of J. Alfred Pfrufrock).
Parismony and Modularity
The term modularity, at least as defined in the dictionary, provides no realistic perspective on the real complexities of the brain. It is a term that emerged from the field of engineering, where it was convenient to design complex circuits so they could be repaired by exchanging large components, as opposed to tracking down problems in the detailed connections of spaghetti-like electronic circuits. Contrary to what PITCHFORD suggests, the term was never common in neurology, even though recently we do find even some of our respected neuroscience colleagues falling into that sloppy usage ever more frequently, now that it has become a meme of massive proportions in so many academic circles. No one wants to be left out of such discussions and all too many have come to casually use the term, even though neuroscience disciplines have had an abundance of apt brain terms (e.g., systems, networks or circuits), long before the modularity idea was introduced to us by individuals who had little direct experience with brain research. This ‘digibit-module’ metaphor (we assume some programmers still fondly remember those useful electronic units prevalent in the 1970s) offers no promising empirical payoffs. Modularity simply adds nothing useful to existing neuroscience strategies or agendas, except perhaps a false sense of security that we have identified a natural kind in the mind/brain without having done any neuroscience. Is it too much to ask all evolutionary psychologists to simply drop the modularity concept? Nothing would be lost and a lot could be gained, as was evident in one recent contribution (DUCHAINE/COSMIDES/TOOBY 2001). If we need such a concept, why not simply use a classic designator such as faculties, which simply helps underscore key problems that needed scientific attention.
We submit that if we do not understand the ancestral minds that are still represented in the many species with which we share the earth, we will not really understand our own. Now that we understand the massive genetic relatedness among all mammals, we must start to fathom how the basic brain processes that all mammals share guide our human mental activities. We suspect that when we begin to accept the existence of certain primitive attentional, emotional and motivational faculties in human mind/brains, we will find some parsimonious ways to solve many of the fascinating problems that evolutionists have brought to the forefront of the psychological agenda. Let us simply share a few examples:
As highlighted at the end of our target article, we believe that human rape emerges substantially from poorly socialized or sociopathic mind/brains that are inclined to harm others for their own primitive sexual gratification. It will be an important neuroscience project to determine to what degree basic aggression systems in the human brain overlap with those that mediate sexual urges. We already know that in certain species the overlap can be substantial (KOLLACK-WALKER/NEWMAN 1995).
As evolutionary psychologists have compellingly shown, males seek beauty and youth in their socio– sexual partners, while females seek signs of devotion and material investment. This may largely be due to the fact that the primitive systems for male erotic urges are driven more by simple exteroceptive cues of nubility, not much different than those of other animals. Is there really much reason to propose any intrinsic cognitive, decision-making circuitry? Might females simply have the socio–neurological skills to respond to primitive social rewards more readily than males—the pleasures of being together, eating together, sleeping together and being tolerant of each other? These are all primitive affective features of the mammalian condition that do not require sophisticated special-purpose, cognitive ‘modules’. This is not to suggest that the origin of such choices are not based on reproductive issues, but simply to
Evolution and Cognition ❘ 73 ❘ 2001, Vol. 7, No. 1
assert that simple emotional systems with a modicum of some general-purpose cognitive skill, may easily yield some of the most striking folk-psychological discoveries of evolutionary psychology.
Yes, childless, rouge males are more likely to harm the young whereas fathers and especially mothers are much less likely to do so, especially to their own (DALY/WILSON 1988). Could this simply reflect the primitive social-bonding mechanisms of the mammalian brain that promote anti-aggressive attitudes by arousing ‘peace-promoting’ neurochemical activities in ancient neural systems? One of the most spectacular findings in this area is the discovery that a single sexual episode can gradually transform infanticidal males into tolerant ones, so that they are least likely to kill young at the time their own offspring enter the world (MENNELLA/MOLTZ 1988). We anticipate that when investigators probe such matters, they will find that satisfying sexual activities arouse the chemistries for nurturant feelings in the human brain, just as they do in other animals (CARTER/LEDERHENDLER/KIRKPATRICK 1997).
We have no problems with the idea that evolutionarily-derived, socio–sexual adaptations exist within our mind/brain dynamics, we simply question the wisdom of conceptualizing these processes as evolved, cortico–cognitive ‘modules’. At the same time, we do agree with carefully analyzing how human cognitive activities change as a function of such affective changes (which will eventually be capable of being precisely manipulated in humans via knowledge derived from the study of the corresponding, emotion-modulating chemistries in animal models). There are other interesting emotional examples to consider from the vantage of the principle of a cross-species parsimony (PANKSEPP 1998a). We certainly believe that such ancient emotional dynamics are bound to have remarkably strong effects on the cognitive dynamics of human minds. However, the study of such processes will require development of a more vigorous, empirically deep psychology (PANKSEPP 1999; SOLMS/NERSESSIAN 1999).
Group Selection
In the initial paper, we chose to leave a variety of our arguments open-ended, so we could attract pointed criticism about issues about which we all truly know very little. The most poignant example was our willingness to consider a viewpoint that is viewed as an anathema by most evolutionists. Namely, the concept of group selection. In truth, we are quite agnostic on this issue and simply urge our colleagues to be
much more open-minded on the topic than many have typically been. First, let us be unambiguous on certain issues. We certainly agree that evolution progresses as a function of the survival of gene patterns. The functional unit of selection must be the individual that carries genes and at least in ‘higher’ animals, that negotiates (typically emotionally) with other members of a species to pass on its genes. In such interactions, affect may be the main currency of decision making and behavioral choice. Thus, it would seem essential to consider the role of emotions, especially those emerging from dyadic interactions, as a fundamental source of evolutionary progression in our species and many others. In short, our introduction of a variant of the group selection concept is only tangentially related to the classic concept.
In its historical meaning, group selection was posed and modeled as a possible explanation of why individuals in social groupings may behave in a way that is overtly suggestive of altruism. WYNN-ED-WARDS (1962) postulated that individuals may concede their own opportunity to pass on their own genetic guideline, by opting to contribute time and resources to other members of the group to which it belonged—a contribution that would facilitate propagation of the species. Soon after, HAMILTON (1964) argued that seemingly altruistic acts could be explained by the concept of inclusive fitness. In this way, individuals would only help out a group member to the degree that the cost of a specific ‘generous’ act was less than the genetic passage to be gained (i.e., the degree of genetic-relatedness should be a predictor of the extent to which an individual will be altruistic). From this vantage, altruism could be explained in terms of the genetic success of individuals, and the potential benefits of reciprocity.
What remains a moot and often an acrimonious point is whether gregarious animals act altruistically when there is negligible genetic gain at stake. In our previous paper, we opined that certain affective/behavioral functions could only propagate within a web of group dynamics. Indeed, if these processes only occur when individuals are genetically related, they certainly can be reduced to selection at the level of the individual. However, if some can also operate strongly between genetically-unrelated individuals (e.g., arising perhaps from pair bonding and other social coalitions) then one can imagine, for instance, dyadic selection (a minimalist form of group selection) to be an influential force in evolution. Either way, we feel that this process is substantially different and thus deserving of its own distinction when
Evolution and Cognition ❘ 74 ❘ 2001, Vol. 7, No. 1
thinking about mechanisms of selection. Henceforth, we will refer to selective processes that operate on neuro/behavioral systems that only function successfully in such groups as social selection (not to be confused with group selection although there is the possibility of linkages).
It is clear that the study of intrinsic social processes was massively marginalized during the previous century. Anyone that has spent time studying such matters should recognize that many socio-affective processes can only operate within groups. Normally, one cannot tickle themselves or socially play alone, and one can only deeply grieve for the loss of another, but not as much for material losses. A preliminary characterization of the underlying brain systems has been achieved for some of these neural processes (PANKSEPP 1998a), but there has been massive resistance in psychological and neuroscience communities for pursuing such topics of ultimate concern. Many still assert that specific social affects do not exist—that they are all learned or socially constructed. This is most certainly wrong, even though all the details of our social existence are surely learned. And when we have the conceptual liberties that the human-type cortex allows, we can obviously choose to go against the dictates of these systems. We are perhaps the only species that has the option of being libertarians.
Our advocacy of the social selection hypothesis was advanced largely to help open scientific inquiry into the many unstudied mysteries that exist in all mammalian brains. We should not close our minds to reasonable possibilities because of the appealing rigor of certain mathematical equations. Whether in some instances social selection and group selection exert a synergistic pressure on certain brain functions remains an intriguing possibility. While it continues to be only a possibility for other species, we feel that it is in fact very plausible that group selection has already occurred in our own species. However, the important distinction is that such selective processes may have been driven primarily by cultural evolution and not by biological evolution (just consider the caste systems in some cultures, not to mention the holocausts that have been inflicted on others). It seems intuitive that gene frequencies and thus collective genomic phenotypes can be altered by seemingly capricious cultural choices. The potential grouping of genes in human beings will be as unpredictable as their new selective force—culture. There should be no reason why this cannot be accepted, without moral exacerbation, as an emergent property of the interactions between dedicated brain
operating systems, flexibly non-dedicated systems (where emergent properties can also be exquisitely housed), environment and culture. It is within such realms where the concept of developmental programs will become most robustly manifest. Cortico– cultural evolution has obviously provided a new and all too often terrible order to the evolved dynamics of the mind/brain (PETERSON 1999), and of course, the genes have also played their part. Fortunately, we still have the hope of better educating our higher cerebral systems.
Behavior Genetics—An Essential Key
It is understandable that evolutionary psychologists did not want to get entangled in the genetic determinism that has historically colored sociobiological thought. However, there is no intellectually-sound reason to be coy about this topic. Genes are obviously overwhelmingly important in the way the mind/brain is constructed as is epigenetic development. To think otherwise is to indulge in politicized nonsense.
The robust heritability of a majority of human personality traits is stunning, while there is very little heritability for the specific things we do in our environments. For years, BOUCHARD (1998) has been finding clear evidence for ‘nature’ in the construction of human personality, while the role for ‘nurture’ has gotten slimmer and slimmer (almost nonexistent by some of his most recent ‘adoptees raised in the same environment’ analyses). Namely, identical twins reared apart show the same reasonably high degree of heritability on practically all traits as do twins reared together (r’s typically between .4 and .6), while two unrelated adoptees raised in the same family exhibit practically zero correlation on such traits (BOUCHARD personal communication with J.P.). Also, it is becoming clear that our genetic dispositions help us actively create social and material environments that are satisfying or less distressing. Yet, no one knows what has really been selected for at the genetic level, but we suggest that at a systems level, it is the vigor of certain basic motivational, emotional and attentional functions. We will not dwell on these important issues since they have been covered so well elsewhere (BOUCHARD 1998), but will share one striking recent example:
BOUCHARD’s most recent work on religiosity was initially a bit of a mess until he broke it down into intrinsic and extrinsic religiosity. The intrinsic variety (e.g., spirituality?) had a high genetic loading, while the extrinsic variety (e.g., church attendance
Evolution and Cognition ❘ 75 ❘ 2001, Vol. 7, No. 1
and other external practices) exhibited a high environmental loading. The two forms of religiousness were inversely correlated—probably not surprising for many evolutionists who understand the utility of deception and the probable importance of cortical processes in generating such tendencies. Also, intrinsic religiosity was positively related to authoritarianism (BOUCHARD et al. 1999). A liberal college education had a small effect on dampening that old time prophet instinct (i.e., a temporary nurture effect), but at eight years past those liberating college days, the old ways were—hang on Evolutionary Psychology fans—coming back with a vengeance, especially if you had lots of kids (personal communication with J.P.). Once again kids force us back to our genetic roots. Clearly, practically any human or animal trait can be amplified or diminished genetically. However, in the past we rarely knew what was really selected, but the new molecular biology will soon change that.
Politics and Tribalism
An enormous amount of ink and mental energy was wasted during the sociobiology wars because of the political inclinations of certain participants (SEGER-STRÅLE 2000). We cannot easily escape our political biases, but intellectual and social disasters can easily be made if we do not own up to our ‘mere beliefs’. For this reason, we especially appreciate SEGER-STRÅLE’s participation in these discussions. For instance, the harsh treatment of the role of genes in our destinies at the hands of politically motivated MARXISTS is a story that presently has only been half told (hopefully, some forthcoming memoirs will set the records straighter). We are the kind of species that can perform intellectual, emotional and bodily monstrosities in behalf of what we deeply believe (PETERSON 1999). All too easily, we develop tribal affiliations which color all our subsequent beliefs and arguments. None of us are immune to such issues, but they are often very apparent when we come to issues as contentious as sociobiology and Evolutionary Psychology (LEWONTIN 1993).
Probably most who read journals such as this are well sensitized (or habituated) to those aspects of the human spirit, but the present debate has certainly not been absent of those features. Let us be so bold as to highlight a striking instance that we could not avoid noticing. Our harshest critic in these discussions, PITCHFORD, asserts that “Neuroevolutionary psychobiology’s reassignment of the tabula rasa to the neocortex revivifies CARTESIAN dualism and represents a desperate strategy likely to be extremely deleterious to scientific investigation. Evolutionary psychology should reject this approach…” To some extent, this assertion is not simply a cold-headed response to the strengths and weaknesses of existing data and arguments, but also an indication of tribal affiliation, since he is a ‘list master’ for the extended evolutionary psychology community. In personal communications, PITCH-FORD was kind enough to say, upon first reading our article, that “although I disagree with a few of the details I do agree with broad thrust of the piece. In my view EP has made a number of fundamental errors: (1) by adopting the method of reverse engineering in place of a consideration of phylogeny (2) by adopting functionalism, or the study of substrate neutral processes, in preference to the study of neurobehavioural and neuropsychological homologies, (3) by adopting a commitment to the notion of the monomorphic mind, thereby excluding a consideration of both polyphenism and adaptive genetic variation as studied by game theorists, and (4) by adopting genic selectionism in preference to developmental systems theory. I think the issue of domain-specific versus domain-general mechanisms is of less importance than these four main issues, as the brain surely contains both, and their relative contributions to our cognitive–emotional capacities is an empirical matter” (personal communication with J.P., 1/3/01).
Then, four months later, after one of us (J.P.) had shared some surprise about the throttling he had provided, PITCHFORD wrote back that “In my view your perspective embraces almost every issue of importance. The commentary does concentrate on areas of disagreement, but had the word limit allowed I would have had much more to say about affective neuroscience as the essential foundation for evolutionary psychology. The emphasis on functionalism that evolutionary psychology has adopted from cognitive science can result in a neglect of cross-species comparisons, and in a reluctance to embrace important branches of evolutionary biology such as ecology and game theory. This can make it seem abiological at times, but I see this as an unwelcome tendency rather than a defining trait. However, most of the faults you identify in ‘seven sins’ can be found in the work of evolutionary psychologists” (personal communication with J.P., 4/23/01). We do not share such personal communications with any desire to embarrass (indeed we agree completely with the gist of those arguments), but they do highlight that in our human (male?) eagerness to put forward strong
Evolution and Cognition ❘ 76 ❘ 2001, Vol. 7, No. 1
views, we often veer far from the middle road. We all tend to do that. We are all prisoners of our times and typically of intellectual tribes that have strong expectations concerning proper behaviors of their members. And all too often we are also captives of our political biases and our remarkable left-hemisphere language skills, which all too often comes close to mere confabulation rather than reasonable argumentation (for perhaps too creative a critique of this human dilemma, see PANKSEPP 2000).
And so what are our own politics? (1) we are committed to accepting our animal nature and at times we may veer rhetorically a bit too far in the zoomorphic direction; (2) we are committed to accepting the affective complexity (and cognitive also) of other animals and we may often sound a bit too anthropomorphic because we do believe that it is fundamentally correct that other mammals have many basic emotional feelings not all that different than our own; (3) we have a commitment to a deep sense of human freedom and plasticity, and we may often claim that humans have more cortico–cognitive choices than they, in fact, do (but we should all hope that they might); and (4) we have a commitment to human justice and paradoxically, that may lead our prose to be a bit more heated than it should be. We truly regret if we were unfair to any of our commentators and we are sorry if some of their important issues were not addressed as fully as they could have been.
The Sin of Anthropocentrism
In closing, let us simply reiterate our thanks to all the commentators for participating in this discussion and refresh our call to the many prominent figures in the field who, so far, have chosen to remain silent on these important issues.1 We are personally committed to the importance of evolutionary views for understanding our deep nature. We do not believe that it can be done without a full confrontation with the emotional tendencies of our fellow animals. We thank evolutionary psycholo-
gists for opening up the debate about human nature once more, and we encourage them to help correct the deep flaw in our educational systems and societal institutions that continue to marginalize the essential understanding of the basic underlying issues—concepts that must be derived from our human understanding but detailed knowledge of which can typically only be derived from well-conducted research on and with our fellow animals. We were quite pleased that GARDNER has found our ‘rat laughter’ research to be intriguing, and that story line continues to yield interesting findings (BURGDORF/ PANKSEPP 2001; BURGDORF et al. 2001).
Our recognition of human potentials will be much more profound when we realize how our general-purpose neocortical expansions have enabled our own capacity to elaborate on the geneticallyingrained solutions that evolution so ‘kindly’ provided. Clearly, our expansive neocortex has permitted the emergence of an unprecedented cultural evolution. This neuro-evolutionary passage emerged too rapidly, or so it seems to us, for any highly-resolved, sociobiologically-engraved strategies to have emerged within the expanding neocortical tissues. Evolutionary psychologists should begin to entertain the full complexity of our animalian nature. With the recent emergence of more balanced views (e.g., BADCOCK 2000), the future of the field seems bright, especially if it can rectify and free itself from the many fledgling mistakes it has already made.
In closing, we also note that many of the issues that we raised in our target article were concurrently being addressed by one of the fathers of modularity and cognitive science. We are delighted that Jerry FODOR recently chose to provide a frank analysis of what has or has not been achieved by the cognitive and evolutionary psychology perspectives: “In fact, what our cognitive science has done so far is mostly to some throw [sic?] light on how much dark there is. So far, what our cognitive science has found out about the mind is mostly that we don’t know how it works” (FODOR 2000, p100). We suggest that this is largely because evolutionary psychology has devoted so little thought to understanding the basic neuro-emotional and motivational processes upon which both human and animal minds are built. This neglect has, we suspect, been largely due to the dualistic, anti-organic bias
Notes
1 We thank all commentators for sharing their perspectives and trust that the thinking of some of the remaining seven investigators who were invited to comment on our ideas have been influenced as well (i.e., David BUSS, Leda COSMIDES, Bobbie LOWE, Randy NESSE, Steven PINKER, John
References
- Badcock, C. (2000) Evolutionary Psychology: A Critical Introduction. Polity Press: Cambridge, UK.
- Baily, P./Davis, E. W. (1943) Effects of lesions of the periaqueductal gray matter in the cat. Proceedings of the Society for Experimental Biology and Medicine 51: 305–306.
- Barkley, R. A. (2001) The excecutive functions and self-regulation: An evolutionary neuropsychological perspective. Neuropsychology Review 11: 1–29.
- Basso, A./Sala, S. D./Farabola, M. (1987) Aphasia arising from purely deep lesions. Cortex 23: 29–44.
- Batro, A. M. (2000) Half a Brain Is Enough: The Story of Nico. Cambridge Univ. Press: Cambridge UK.
- Bauman, M. L./Kemper, T. L. (1995) Neuroanatomical observations of the brain in autism. In: Panksepp, J. (ed) Advances in Biological Psychiatry, Vol. 1, Greenwich CT: JAI Press, pp. 1–26.
- Bechtereva, N. P./Abdullaeva, Y. G. (2000) Depth electrodes in clinical neuropsychology: neuronal activity and human cognition function. International Journal of Psychophysiology 37: 11–29.
- Berridge, K. C. (2000) Measuring hedonic impact in animals and infants: microstructure of affective tast reactivity patterns. Neuroscience and Biobehavioral Reviews 24: 173– 198.
- Berridge, K. C./Valenstein, E. S. (1991) What psychological process mediates feeding evoked by electrical stimulation of the alteral hypothalamus? Behavioral Neuroscience 105: 3–14
- Bouchard, T. J. (1998) Genetic and environmental influences on adult intelligence and special mental abilities. Human Biology 70: 257–279.
- Bouchard, T. J./McGue, M./Lykken, D./ Tellegen, A. (1999) Intrinsic and extrinsic religiousness: Genetic and environmental influences and personality correlates. Twin Research 2: 88–98.
- Buchner, R. L./Corbetta, M./Schatz, J./Raichle, M. S./Petersen, S. E. (1996) Preserved speech abilities and compensation following prefrontal damage. Proceedings of the national Academy of Sciences 93: 1249–1253.
TOOBY and E. O. WILSON). The editors are willing to consider sharing their comments in future issues of this journal.
- 2 We typically use Evolutionary Psychology in capitals when we refer to the revolutionary, social-psychological version of the discipline, and lower case when we refer to the more generic variety.
- Borod, J. C. (2000) The Neuropsychology of Emotion. Oxford University Press: New York.
- Breitenstein, C./Van Lancker, D./Daum, I./Waters, C. H. (2001) Impaired perception of vocal emotions in Parkinson’s disease: Influence of speech time processing and executive functioning. Brain and Cognition 45: 277–314.
- Burgdorf, J./Knutson, B./Panksepp, J./Ikemoto, S. (2001) Nucleus accumbens amphetamine microinjections unconditionally elicit 50 kHz ultrasonic vocalizations in rats. Behavioral Neuroscience. In Press.
- Burgdorf, J./Knutson, B./Panksepp, J./Shippenberg, T. (2001) Ultrasonic vocalizations index pharmacological aversion in adult rats. Psychopharmacology 155: 35–42.
- Burgdorf, J./Panksepp, J. (2001) Tickling induces reward in adolescent rats. Physiology & Behavior 72: 167–173.
- Carter, S./Kirkpatrick, B./Lederhendler, I. I. (eds) (1997) Neurobiology of affiliation. Annals of the New York Academy of Sciences 807. New York Academy of Sciences: New York.
- Castro-Caldas, A./Petersson, K. M./Reis, A./Stone-Lander, S./ Ingvar, M. (1998) Learning to read and write during childhood influences the functional organization of the adult brain. Brain 121: 1053–1063.
- Charney, D. S./Nestler, E. J./Bunney, B. S. (eds) (1999) Neurobiology of Mental Illness. Oxford University Press: New York.
- Cosmides, L./Tooby, J. (2000) Evolutionary psychology and the Emotions. In: Lewis, M./Haviland, J. (eds) The Handbook of Emotions, 2nd edition. Guilford: New York, pp. 91–116.
- Crosson, B./Zawacki, T./Brinson, G./Lu, L./Sadek, J. R. (1997) Models of subcortical functions in language: current status. Journal of Neurolinguistics 10(4): 277–300.
- Daly, M./Wilson, M. (1988) Homicide. Aldine: New York.
- Damasio, A. R. (1999) The Feeling of What Happens, Body and Emotion in the Making of Consciousness. Harcourt Brace: New York.
- Darwin, C. (1998) The Expression of the Emotions in Man and Animals, 3rd edition. Oxford University Press: New York. Originally published 1872.
- Dawson, G./Fisher, F. W. (eds) (1984) Human behavior and
Evolution and Cognition ❘ 78 ❘ 2001, Vol. 7, No. 1
the developing brain. Guilford Press: New York.
de Waal, F. (2001) The Ape and the Sushi Master. Basic Books: New York.
- Dennet, D. (1995) Darwin’s Dangerous Idea. Simon and Schuster: New York.
- Duchaine,B./Cosmides,L./Tooby,J. (2001) Evolutionary psychology and the brain. Current Opinions in Neurobiology 11: 225–230.
- Dugatkin, L. A. (2000) The Imitation Factor. The Free Press: New York.
- Feinberg, T. E. (2001) Altered Egos: How the Brain Creates the Self. Oxford Univ. Press: New York.
- Fifer, W. P./Moon, C. M. (1994) The role of mother’s voice in the organization of brain functions in the newborn. Acta Paediatrica 83: 86–93.
- Freeman, W. J. (1999) How the brain makes up its mind. Blackwell: London UK.
- Fodor, J. (2000) The Mind Doesn’t Work That Way, The Scope and Limits of Computational Psychology. The MIT Press: Cambridge UK.
- Gallese, V./Goldman, A. (1998) Mirror neurons and the simulated theory of mind-reading. Trends in Congnitive Sciences 2: 493–501.
- Gallistel, C. R./Gibbon, J. (2001) The Symbolic Foundations of Conditioned Behavior. Lawrence Erlbaum Assocs: Mahwah NJ.
- Griffiths, P. E. (1997) What Emotions really Are: The Problem of Psychological Categories. University of Chicago Press: Chicago IL.
- Grabowski, T. J./Damasio, A. R. (2000) Investigating language with functional neuroimaging. In: Toga, A. W./ Mazziotta, J. C. (eds) Brain Mapping: The Systems. San Diego CA: Academic Press, pp. 425–461.
- Hamilton, W. D. (1964) The genetical evolution of social behaviour (parts I and II). Journal of Theoretical Biology 7: 1– 52.
- Ikemoto, S./Panksepp, J. (1999) The role of nucleus accumbens dopamine in motivated behavior: A unifying interpretation with special reference to reward-seeking. Brain Research Reviews 31: 6–41.
- Jacobs, B./Scheibel, A. B. (1993) A quantitative dendritic analysis of Wernicke’s are in humans. I. Lifespan changes. Journal of Comparative Neurology 327: 83–96.
- Jacobs, B./Schall, M./Scheibel, A. B. (1993) A quantitative dendritic analysis of Wernicke’s area in humans: II. Gender, hemispheric, and environmental factors. Journal of Comparative Neurology 327: 97–111.
- John, R./Surani, M. (1996) Imprinted genes and regulation of gene expression by epigenetic inheritance. Current Opinion in Cell Biology 8: 348–353.
- Kaplan-Solms, K./Solms, M. (2000) Clinical Studies in Neuro-Psychoanalysis, Introduction to a Depth Neuropsychology. Karnac Books: London UK.
- Kim, J. (1993) Supervenience and Mind: Selected Philosophical Essays. Cambridge University Press: New York.
- Keverne, E. B./Fundele, R. et al. (1996) Genomic Imprinting and the Differential Roles of Parental Genomes in Brain Development. Developmental Brain Research 92: 91–100.
- Kollack-Walker, S. S./Newman, S. W. (1995) Mating and agonistic behavior produce different patterns of Fos immunolabeling in the male Syrian hamster. Neuroscience 14: 721–736.
- Lane, R. D./Nadel, L. (eds) (2000) Cognitive Neuroscience of Emotion. Oxford University Press: New York.
- Lander, E. S. et al. (2001) Initial sequencing and analysis of the human genome. Nature 409: 860–921.
- Lesch, K. P./Bengel, D./Heils, A./Sabol, S. Z./Greenberg, B. D./Petri, S./Benjamin, J./Müller, C. R./Hamer, D. H./Murphy, D. L. (1996) Association of anxiety-related traits with a polymorphism in the serotonin transporter gene regulatory region. Science 274: 1527–1531.
- Lesch, K. P./Merschdorf, U. (2000) Impulsivity, aggression, and serotonin: a molecular psychobiological perspective. Behavioral Sciences and the Law 18: 581–604.
- Lewontin, R (1993) Biology as ideology: the doctrine of DNA. Harper Perennial: New York.
- Lieberman, P. (2000) Human Language and Our Reptilian Brain: The Subcortical Bases of Speech, Syntax and Thought. Harvard Univ. Press: Cambridge MA.
- Lucki, I. (1998) The spectrum of behaviors influenced by serotonin. Biological Psychiatry 44: 151–162.
- Mennella, J./Moltz, H. (1988) Infanticide in rats: Male strategy and female counter-strategy. Physiology & Behavior 42: 19–28.
- Nestler, E. J. (1998) Antidepressant treatments in the 21st century. Biological Psychiatry 44: 526–533.
- Neville, H. J./Bavelier, D./Corina, D./Rauschecker, J./Karni, A./Lalwani, A./Braun, A., Clark, V., Jezzard, P. & Turner, R. (1998) Cerebral organization for language in deaf and hearing subjects: Biological constraints and the effects of experience. Proceeding of the National Academy of Sciences 95: 922–927.
- Nimchinsky, E. A./Gilissen, E./Allman, J. M./Perl, D. P./Erwin, J. M./Hof, P. R. (1999) A neuronal morphologic type unique to humans and great apes. Proceedings of the National Academy of Sciences 96: 5268–5273.
- Nudo, R. J./Wise, B. M./SiFuentes, F./Milliken, G. W. (1996) Neural substrate for the effects of rehabilitative training on motor recovery after ischemic infarct. Science 272: 1791– 1794.
- Oyama, S. (1985) The Ontogeny of Information: Developmental Systems and Evolution. Cambridge UK: Cambridge University Press. Rreprinted in 2000 by Duke University Press.
- Panksepp, J. (1981) Brain opioids: a neurochemical substrate for narcotic and social dependence. In: Cooper, S. (ed) Progress in Theory in Psychopharmacology. Academic Press: London, pp. 149–175.
- Panksepp, J. (1982) Toward a general psychobiological theory of emotions. The Behavioral and Brain Sciences 5: 407– 467.
- Panksepp, J. (1986) The neurochemistry of behavior. Annual Review of Psychology 37: 77–107.
- Panksepp, J. (1988) Brain emotional circuits and psychopathologies. In: Clynes, M./Panksepp, J. (eds) Emotions and Psychopathology. Plenum Press: New York, pp. 37–76.
- Panksepp, J. (1989) The psychobiology of emotions: The animal side of human feelings. In: Gainotti, G./Caltagirone, C. (eds) Emotions and the dual brain. Experimental Brain Research Series 18. Springer-Verlag: Berlin, pp. 31–55.
- Panksepp, J. (1990) The psychoneurology of fear: evolutionary perspectives and the role of animal models in understanding anxiety. In: Burrows, G. D./Roth, M./Noyes Jr., R. (eds) Handbook of Anxiety, Vol. 3: The Neurobiology of Anxiety. Elsevier: Amsterdam, pp. 3–58.
- Panksepp, J. (1993) Neurochemical control of moods and emotions, amino acids to neuropeptides. In: Lewis, M./ Haviland, J. (eds) The Handbook of Emotions. Guilford: New York, pp. 87–107.
- Panksepp, J. (1998a) Affective Neuroscience, The Foundations of Human and Animal Emotion. Oxford University Press: New York.
Evolution and Cognition ❘ 79 ❘ 2001, Vol. 7, No. 1
- Panksepp, J. (1998b) Attention deficit disorders, psychostimulants, and intolerance of childhood playfulness: A tragedy in the making? Current Directions in psychological Sciences 7: 91–98.
- Panksepp, J. (1998c) The quest for long-term health and happiness: To play or not to play, that is the question. Psychological Inquiry 9: 56–65.
- Panksepp, J. (1999) Emotions as viewed by psychoanalysis and neuroscience: An exercise in consilience. Neuro-Psychoanalysis 1: 15–38.
- Panksepp, J. (2000a) The neurodynamics of emotions: an evolutionary-neurodevelopmental view. In: Lewis, M. D./ Granic, I. (eds) Emotion, Self-Organization, and Development. Cambridge Univ. Press: New York, pp. 236–264.
- Panksepp, J. (2000b) Affective consciousness and the instinctual motor system. The neural sources of sadness and joy. In: Ellis, R./Newton, N. (eds) The Caldron of Consciousness, Motivation, Affect and Self-organization. Vol 16. Advances in Consciousness Research. John Benjamins Pub. Co: Amsterdam, pp. 27–54.
- Panksepp, J. (2000c) On preventing another century of misunderstanding, Toward a psychoethology of human experience and a psychoneurology of affect. Neuro-Psychoanalysis 2: 240–255.
- Panksepp, J. (2001a) The long-term psychobiological consequences of infant emotions: Prescriptions for the twentyfirst century. Infant Mental Health Journal 22: 132–173.
- Panksepp, J. (2001b) At the interface between the affective, behavioral and cognitive neurosciences: Decoding the emotional feelings of the brain. Brain and Cognition, in press.
- Panksepp, J./Normansell, L./Cox, J. F./Siviy, S. M. (1994) Effects of neonatal decortication of social play of juvenile rats. Physiology & Behavior 4: 29–443.
- Panksepp, J./Miller, A. (1996) Emotions and the aging brain: regrets and remedies. In: Magai, C./McFadden, S. H. (eds) Handbook of Emotions and Aging. Academic Press: New York, pp. 3–26.
- Panksepp, J./Knutson, B./Pruitt, D. L. (1998) Toward a neuroscience of emotions: the epigenetic foundations of emotional development. In: Mascolo, M. F./Griffin, S. (eds) What develops in emotional development? Plenum Press: New York, pp. 53–84.
- Pasqual-Leone, A./Torres, F. (1993) Plasticity of the sensorimotor cortex representations of the reading finger in Braille readers. Brain 116: 39–52.
- Peterson, J. B. (1999) Maps of Meaning: The Architecture of Belief. Routledge: New York.
- Rilling, J. K./Insel, T. R. (1999) The primate neocortex in comparative perspective using magnetic resonance imaging. Journal of Human Evolution 37: 191–223.
- Rinn, W. E. (1984) The neuropsychology of facial expression, A review of the neurological and psychological mechanisms for producing facial expressions. Psychological Bul-
letin 95: 52–77.
- Rizzolatti, G./Fadiga, L./Gallese, V./Fogassi, L. (1996) Premotor cortex and the recognition of motor actions. Brain Research and Cognition 3: 131–141.
- Robinson, T. E./Berridge, K. C. (1993) The neural basis of drug craving: an incentive-sensitization theory of addiction. Brain Research and Reviews 18: 247–291.
- Segerstråle, U. (2000) Defenders of the Truth, The Battle for Science in the Sociobiology Debate and Beyond. Oxford University Press: New York.
- Semendeferi, K./Damasio, H. (2000) The brain and its main anatomical subdivisions in living hominoids using magnetic resonance imaging. Journal of Human Evolution 38: 317–332.
- Shand, A. F. (1920) The Foundations of Character, Being a Study of the Tendencies of the Emotions and Sentiments. MacMillan and Co: London UK.
- Schiff, N. D./Plum, F. (1999) The neurology of impaired consciousness: global disorders and implied models. ASSC Eseminar target article. Retrieved on Dec 2, 1999, from the World Wide Web: http://www.phil.vt.edu/assc/niko.html.
- Solms, M. (1997) The Neuropsychology of Dreams: A Clinico-Anatomical Study. Lawrence Erlbaum Associates: New York.
- Solms, M./Nersessian, E. (1999) Freud’s theory of affect, questions for neuroscience. Neuro-Psychoanalysis 1: 5–14.
- Steiner, J. E./Glaser, D./Hawilo, M. E./Berridge, K. E. (2001) Comparative expression of hedonic imact: affective reactions to taste by human infants and other primates. Neuroscience and Biobehavioral Reviews 25: 53–74.
- Thatcher, R. W./Reid Lyon, G./Rumsey, J./Krasnegor, N. (eds) (1996) Developmental Neuroimaging: Mapping the Development of Brain and Behavior. Academic Press: San Diego.
- Toga, A. W./Mazziotta, J. C. (2000) Brain Mapping: The Systems. Academic Press: San Diego CA.
- Valenstein, E. S. (1998) Blaming the brain. The Free Press: New York.
- Valenstein, E. S./Cox, V. C./Kakolewski, J. W. (1970) Reexamination of the role of the hypothalamus in motivation. Psychological Review 77: 16–31
- Vargha-Khadem, F./Carr, L. J./Isaacs, E./ Brett, E./Adams, C./ Mishkin, M. (1997) Onset of speech after left hemispherectomy in a nine-year-old boy. Brain 120: 159–182.
- Venter, J. C. et al. (2001) The sequence of the human genome. Science 291: 1304–1351.
- Wallin, N. L./Merker, B./Brown, S. (1999) The origins of music. MIT Press: Cambridge MA.
- Wimmer, M./Ciompi, L. (1995) Evolutionary aspects of affective-cognitive interactions in the light of Ciompi’s concept of ‘affect-logic’. Evolution and Cognition 2(1): 37–58.
- Wynn-Edwards, V. C. (1962) Animal Dispersion in Relation to Social Behavior. Oliver & Boyd: Edinburg.
An Evolutionary Perspective on Cognitive Orientation
Introduction
The purpose of this paper is twofold. The first goal is to describe how the cognitive guidance of behavior has developed in an evolutionary sense. The conception that cognition plays a role in regard to survival and behavior lies at the core of the evolutionary epistemology theories that consider knowledge as an evolutionary process subserving survival (CAMPBELL 1974; LORENZ 1977; POPPER 1984; RIEDL 1980). These theories provide the bioethological basis for the scientific endeavor to describe how cognition guides behavior. Starting
Abstract
The paper deals with evolutionary aspects of the cognitive guidance of behavior. It is assumed that cognition guides behavior because of empirical evidence and because the impressive development of cognition in the course of evolution suggests that it plays a role in regard to survival. The theory of cognitive orientation—a theory of cognitive motivation on the human level—is presented as a basic framework for the evolutionary analysis. The major advantages of this theory are that it provides a comprehensive description of the processes intervening between input and output and has ample empirical support. The major evolutionary aspects discussed concern the differentiation between motivation and cognition, the development of meaning and the development of action, emphasizing the role of cognition in regard to each.
Key words
Cognition, development, meaning, belief, behavioral program.
with a cognitive theory of motivation on the human level, we will examine its evolutionary development and implications. In view of the novelty of the goal, that which we can expect to do in a first exploratory paper of this kind is to confront the issue of the evolutionary aspects of the cognitive guidance of behavior and trace some of the major tracks along which the cognitive guidance of behavior seems to have developed in the framework of evolution. In order to clarify our question, it is advisable to emphasize that while it is well-known that in the course of evolution both cognition and behavior have developed each in its own right and in terms of its own characteristics, it is neither well-known nor obvious that the cognitive guidance of behavior did. This is precisely our thesis. Starting from the assumption that in living organisms cognition guides behavior (KREITLER/KRE-
erties. Thus, our exploration will be guided by questions, such as “What happened to the cognitive guidance of behavior in the course of evolution?”, “What has evolution done to it?”, “How was it affected by evolution?”, “Which of its components were shaped and transformed by evolution?”
Our second goal has to do with SKINNER. Years ago during a Sabbatical stay at Harvard University I had the chance to discuss psychological issues with SKIN-NER, who at the time was already retired from Harvard University (1975–1976). As it happened I was lucky enough to have stayed in the same neighborhood in Cambridge, MA where he lived and thus had ample opportunity of discussing science with him. After he became familiar with the theory of the cognitive guidance of behavior that Hans KREITLER and I had developed—the theory of cognitive orienta-
Evolution and Cognition ❘ 81 ❘ 2001, Vol. 7, No. 1
ITLER 1976), the question of the evolutionary development of the cognitive guidance of behavior appears to be both legitimate and relevant. It is legitimate because any phenomenon, structure or process that has been observed on the human level has analogues or primary rudimentary manifestations on earlier evolutionary levels (MITHEN 1996; THORPE 1974). It is relevant because considering the evolutionary understructure and aspects of a basic human process provides increased insights and opportunities for deeper understanding of its structural and functional prop-
Shulamith Kreitler
tion—he said one day (I bring his views in a free paraphrase of my notes from the time): “The cognitive orientation theory is the extreme opposite of the brand of behaviorism that I have developed and which came to be called SKINNERIANISM. It is opposite because it starts on the top, uses an apparently unparsimonious set of theoretical concepts, assumes primacy of the cognitive system, and attributes a central role to cognition in the functioning of organisms. In contrast, SKINNERIANISM starts at the very bottom, uses an apparently monastically frugal set of a-theoretical concepts, does not accord any status to the cognitive system, whose existence it barely acknowledges and considers as a theoretical outrage, and attributes no role to it in the functioning of organisms. However, an integral part of the development of an approach in science (viz. theory) is to test its extensibility. Therefore, I was intent to test how far up the evolutionary ladder I can get, in order to find out how much I can explain with my theoretically meager tools on the higher levels of psycho– biological functioning. That’s why I tried to explain even politics in behaviorist terms and why I wrote Walden Two, a book about a utopian society based only on operational conditioning. You should do the same with the cognitive orientation theory, though in an inverse sense. Your task is to test how far down the evolutionary ladder you can get with the cognitive orientation theory, in order to find out how much you can explain with your theoretically rich tools on the lower levels of psycho–biological functioning preceding the human. Since you start with a big load of concepts, stepping down the evolutionary ladder should give you a chance to see which components developed and why. Frankly, I envy you this task”. So, this is the task with which this paper deals. Notably, the procedure suggested by SKINNER corresponds to the use of evolutionary thinking within the top-down system advocated by DEKAY and SHACKELFORD (2000). Handling this task was delayed so much probably because we had first to learn much more about the cognitive orientation theory before we could put it to the test of ‘extensibility’.
Cognition: What It Is not and What It Is
The basic thesis of the cognitive orientation (CO) theory is that cognitive contents and processes guide behavior. Cognitive contents refer to information, beliefs, attitudes, opinions, values, and meanings in the broad sense of the term, whereas cognitive processes are operations and transformations of different kinds that deal with the production, testing, elaboration, transformation, analysis, storing, retrieval and uses of cognitive contents. Cognition, as defined above, is not restricted to any particular species and occurs on different evolutionary levels. In principle, we should expect its barest primary manifestations in the simplest organisms (RIEDL 1980, 1981, 1987).
In order to understand better the role of cognition in the motivational context, it is necessary to emphasize that cognition does not entail rationality and not even reasonableness. The fact that humans may think rationally does not mean that they always or mostly do or that this kind of thinking prevails in the motivational context. Rationality constitutes one set of rules which may guide cognition, but there are other sets, such as Aristotelian logic, Boolean logic, mythological thinking, symbolic thinking, magical thinking etc. (CASSIRER 1953; KREITLER 1965; LÉVI-STRAUSS 1979).
Likewise, the involvement of cognition in motivation does not mean that motivation depends on or involves a decisional process which considers, for example, benefits and costs, advantages and disadvantages, pros and cons, weighing each carefully and calculating the end-result according to normative decision rules (as some models of cognitive motivation assume, e.g., BECKER 1974; ROGERS 1983; RONIS 1992).
Further, cognition does not entail consciousness, nor is it a product or function of consciousness. Consciousness is itself a function of cognition, and may affect cognitive functioning but is not a necessary condition for cognition (KREITLER 1999). Cognitive processes run their course without consciousness as most other organismic processes. They may become conscious to a certain extent, under specific conditions, when this subserves the needs of the organism.
Similarly, cognition does not entail voluntary control. Though the exercise of so-called voluntary control involves cognition, the involvement of cognition does not mean that one has voluntary control over any aspect of the motivational dynamics of behavior.
Finally, cognition is not restricted to the verbal medium. Indeed, cognitive processes may occur without relying on any formal medium of expression, and when some of their aspects or end-results are expressed or communicated this may occur by means of any form of expression, verbal or nonverbal.
So, given that cognition is not necessarily logical, reasonable, conscious, voluntarily controlled and verbal, which positive property can be attributed to it? We venture to suggest that it is meaning-dependent.
Evolution and Cognition ❘ 82 ❘ 2001, Vol. 7, No. 1
Meaning is a concept that plays a major role in cognition. It is defined as a pattern of cognitive contents (called ‘meaning value’, e.g., a color, an emotion) focused on some input (called referent, e.g., a stimulus, an object) that is expressed verbally or nonverbally, and forms together with the input or subject a meaning unit. Examples of meaning units are The sea—blue; Table—made of wood; Car—with four doors; Virus—scary. In a given context meaning consists of a sequence of meaning units, each of which may be characterized in terms of the contents assigned to the subject (e.g., causes, sensory qualities) and the relations between it and the subject (e.g., its directness, generality). The characterizations constitute a set of variables (called ‘meaning variables’) which enable assessing the meaning of any input or the meaning assignment tendencies of an individual. Meaning includes the more interpersonal part as well as the more personal–subjective part, and may vary in contents, structure, variety and complexity (KREITLER/KREITLER 1985, 1986, 1987a, 1989, 1990a, 1990b; KREITLER 1999).
Meaning provides the raw materials for the contents of cognition and the functional frames for the processes of cognition. Accordingly, cognition may be defined as a system which produces, elaborates, transforms, stores and uses meanings. Hence, the properties, contents, processes, functioning and products of cognition are functions of meaning.
When we conceive of a cognitive model of motivation, we talk in fact of a model of motivation in which meaning fulfills a major role. Basically, the CO model is a model based largely—though not exclusively—on the construction, transformation and application of meanings. It describes the processes intervening between input and output not as a series of rational decisions or problem solving, but as outputs of meanings and clustered orientative beliefs, which give rise to motivational dispositions manifested in behavior.
The Cognitive Orientation Theory: General Characteristics
In its original presentations CO was a theory of cognitive motivation of behavior elaborated in regard to overt behaviors (KREITLER/KREITLER 1965, 1972, 1976, 1982). Later developments led to the elaboration of models applicable to further specific domains, such as emotions, physiological processes of disease and health, psychopathology and cognitive behaviors (KREITLER 1999; KREITLER/KREITLER 1986, 1989, 1991a, b, 1997). Accordingly, at present it would be justified to distinguish between the different more specific CO models and the comprehensive CO theory, which refers to the general theoretical concepts and procedures characteristic for all these models. In the present context we will focus on the basic CO theory and CO model of behavior.
CO is a cognitive model of behavior designed to provide an account of the major processes intervening between input and output and to enable understanding, predicting and changing human behavior. This introductory statement emphasizes several important characteristics of the CO model. First, as noted, the original application domain of CO has been human behavior. Secondly, it is a model focused on actual observable overt behavior, as distinguished from self-reported behavior or decisions to act, commitments to action and other replacements of behavior. Thirdly, the purpose of the model is to provide concepts and methods that would be directly applicable for understanding retrospectively, predicting prospectively, and changing at present human behavior. While different psychological methods allow in principle indirectly the attainment of the specified goals, the special characteristic of the CO model is that it enables attaining the mentioned goals by the same methods (rather than one goal by one method, and another goal by another method) and that it functions by direct application of the methods so that interpretational pitfalls are avoided on the way. Fourthly, the CO model is a comprehensive model that addresses all the major links and stages intervening between the input and the output, in contrast to other motivational models that focus on one or another stage or aspect of the process (e.g., planned behavior, AJZEN 1985; health belief model, BECKER 1974).
The basic thesis of the CO theory is that cognitive contents and processes guide behavior. The rationale for this thesis is twofold. The first reason is rooted in the empirical–scientific approach of psychology. On the human level there is a large body of research demonstrating the contributions of cognition to behavior and the utility of this assumption (for a review see KREITLER 2002).
The second reason is rooted in the more general evolutionary approach. It assumes that since cognition has undergone such a spectacular development in the course of evolution (WILSON 1978, p87), it is hardly possible that it does not contribute to behavior which is the prime tool of survival. In this sense, CO is embedded in evolutionary epistemology, which conceives of a continuum from biological to
Evolution and Cognition ❘ 83 ❘ 2001, Vol. 7, No. 1
cognitive processes and attributes to cognition a role in survival (KOVÁ 2000; LORENZ 1977; PLOTKIN 1997; RIEDL 1980, 1987). Cˇ
Accordingly, CO is a behavior-oriented theory, which means that whatever the role of cognition, CO is not a theory of cognition but of motivation and the focus is always on behavior. The purpose of applying cognition is to understand, describe, predict or change behavior.
In addition to the basic thesis about the involvement of cognition in motivation, the CO theory assumes that there is a basic distinction between the motivational disposition for behavior and the performance of behavior. Both are indispensable for any act, at least on the human level. The disposition provides the directionality and answers, as it were, the questions “Why did this behavior occur?” or “Why did the behavior occur specifically in this direction?” The performance of behavior is the operational manifestation of the directionality and answers, as it were, questions, such as “Why did behavior take the form that it did?”, “Why was the behavior executed specifically in this manner or with these means?” The distinction between the directionality of behavior and the manner of its enactment is evident also on lower evolutionary levels, even in regard to reflexes, defensive responses and tropisms. Referring, for example, to the flight of bees, GALLISTEL (1990, pp527–528) writes: “The behavioral purpose is established by the bee’s motivational state, which is conceived of as a set of signals that selectively potentiates a functionally cohesive set of acts” (see ibid also pp87–88 concerning the migration of birds). The distinction itself, its sharpness and manifestations are an evolutionary product (see “Evolutionary aspects of the CO theory”).
The motivational disposition and the performance of behavior differ greatly in their components, structure, and dynamics. Also, the involvement of cognition in motivation differs in regard to the disposition and the performance, as will become evident in the next section.
The last general characteristic of the CO theory to be mentioned in this context concerns the multileveledness of its structure and functioning. Rather than assuming a dominant single course of action on a relatively high level, according to the CO model the input is first addressed in terms of the lower and simpler levels of functioning, that are replaced by more complex ones only if they fail to deal with the situation, according to concurrent or previous experience. The lower and simpler levels of functioning correspond to those that may be identified in lower evolutionary levels. They persist in more advanced species, such as the human, possibly in order to handle simpler situations or more basic survival tasks. Be it as it may, the persistence of the lower levels side by side with the higher levels as functional options within the same organism are in accord with the basic evolutionary principle whereby higher developmental products do not necessarily displace or replace lower ones, but may rather complement, transform and limit their scope. An example is provided by the different stages of evolution contained within the brain of homo sapiens, sometimes called the reptilian, mammalian and human brain (MCLEAN 1973). This phenomenon may be called “preservation of the fittest”. Moreover, the CO theory assumes that issues are handled first on the lower levels, or alternately, that whatever can be handled on a lower level is not handled on a higher level. This set of principles built into the CO model demonstrates the extent to which the evolutionary perspective has been built into the theory down to its fundamentals.
The CO Theory: How Does It Function? Major Theoretical Stages
In the present context only a brief presentation of the CO model of behavior is provided. It will be described in its original form, as it applies on the human level. Later sections will focus on evolutionary aspects.
The CO theory provides detailed descriptions of the processes intervening between input and output. These can be grouped into four stages, each characterized by metaphorical questions and answers. The first stage is initiated by an external or internal input and is focused on the question “What is it?” It consists in identifying the input in terms of a limited and primary ‘initial meaning’ as one of the following: (a) a signal for a defensive, adaptive or conditioned response, or (b) a signal for molar action, or (c) as irrelevant in the present situation, or (d) as new or especially significant, and hence as a signal for an orienting response.
The first alternative consists in the evocation of a defensive or adaptive response, unconditioned or conditioned. If feedback indicates that the input has been properly handled, the behavioral act is brought to an end. The second alternative consists in the emergence of a meaning that identifies the input out from the start as one that cannot be handled by means of a conditioned or unconditioned response and calls for a response on a higher level (viz. molar), that requires more preparatory elaboration. The
Evolution and Cognition ❘ 84 ❘ 2001, Vol. 7, No. 1
third alternative consists in the emergence of a meaning that includes no components enabling, indicating or compelling for action. Hence, the input is dismissed as irrelevant. The fourth alternative arises when the input cannot be identified, for example, because too little information is available, or no resemblance could be established between it and patterns of stimuli stored in memory. Hence, it is transferred to a higher level of elaboration.
The second stage is initiated by feedback indicating failure of the conditioned or unconditioned responses to cope with the situation, or by a meaning signalling the need for molar action, or by an input that has failed to be sufficiently identified despite the orienting response. It is focused on the question “What does it mean in general and what does it mean to or for me?” An enriched process of meaning generation sets in, based on extended elaboration of both the interpersonally-shared and the personal kinds of meaning, in terms of meaning units in the form of beliefs. Beliefs are cognitive units (namely, they include a referent and a meaning value), expressed verbally or not, which express some message presented as true (acceptable, adequate, etc.) or not. The beliefs that emerge at this stage may refer to the individual’s goals, norms, oneself, or others and reality. By examining the extent to which the person’s goals, norms, beliefs about oneself and reality are involved, meaning generation leads eventually to a specification whether action is required or not. Insofar as meaning elaboration indicates that the situation is relevant in regard to the person’s goals, norms, beliefs about self or general beliefs, the indication is pro-action or at least, going on with the process of elaborating the input, even though the nature of the action or its direction remain unspecified. In this context, relevance of beliefs means that a goal arises which may seem to be attainable in the situation, a norm belief is evoked which implies the necessity of undertaking action, or a belief about oneself or a general belief arises which indicates possible dangers in the situation if no action is undertaken or if action is undertaken. Thus, the output of the second stage is indication for action, blocking of action, or no indication for action.
If there is indication for action or at least no blocking of action, the third stage sets in. It is focused on the question “What will I do?” The answer is sought by means of beliefs characterized in terms of relevant contents and a specific form. Relevance in contents denotes deep underlying meanings of the involved inputs rather than the superficial obvious explicit meanings. The deep meanings are characterized
| Belief | Subject | Relation |
|---|---|---|
| Self | I | Factual |
| Norms | Non-I | Desirable |
| Goals | I | Desired |
| General | Non-I | Factual |
Figure 1: Structure of the Four Types of Beliefs.
first, by being of the personal–subjective type rather than the interpersonally-shared type and second, by being relatively remote though not dissociated from the surface meanings. For example, when the input is ‘dieting’, the surface meaning is likely to be “eating less”, and the deeper personal meanings, which do not necessarily refer specifically to eating, could be “renouncing pleasures”, or “accepting restrictions on one’s actions”. Meaning elaboration of the input in successive steps leads to the emergence of different deeper meanings.
As noted, the beliefs are characterized also in terms of form. The beliefs that play a role in the third stage are formally of four types: (a) Beliefs about goals, which express actions or states desired or undesired by the individual (e.g., “I want to be respected by others”); (b) Beliefs about rules and norms, which express ethical, esthetic, social and other rules and standards (e.g., “One should be assertive”); (c) Beliefs about self, which express information about oneself, such as one’s habits, actions, feelings or abilities (e.g., “I rarely become excited”, “I was born in Israel”); and (d) General beliefs, which express information concerning others and the environment (e.g., “The world is a highly dangerous place”). The four types of beliefs may be characterized formally in terms of the kind of subject and the kind of relation between the subject and predicate. Figure 1 presents the formal structure of the four types of beliefs.
Accordingly, the cognitive elaborations in the third stage refer to beliefs of four types that represent deep underlying meanings of the involved inputs. The meaning elaborations consist in matchings and interactions between beliefs (‘belief clustering’), based on clarifying the orientativeness of the beliefs (i.e., the extent to which they support the indicated course of action). If the majority of beliefs of a certain type support the action, that belief type is considered as positively oriented in regard to that action. Alternately, it may be negatively oriented or lack any orientativeness. If all four belief types support a certain action, or at least three support it whereby the fourth
Evolution and Cognition ❘ 85 ❘ 2001, Vol. 7, No. 1
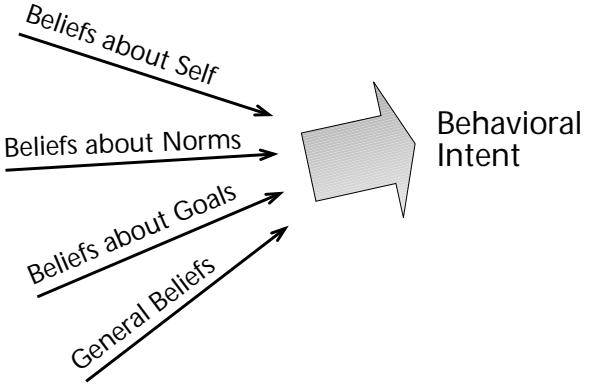
Figure 2: The Behavioral Intent.
is neutral, a cluster of beliefs is formed (‘CO cluster’) orienting toward a particular act. It generates a unified tendency orienting toward the performance of an action, which is called behavioral intent, and can be considered as a vector representing the motivational disposition towards a given behavior (see Figure 2).
In many cases the process of forming a CO cluster is greatly reduced because there exist in memory almost complete CO clusters, formed on the basis of past recurrences of behaviors in similar situations. The meanings of the input allow for retrieving such almost-complete CO clusters and adapting them to the present situation by the addition or transformation of a few beliefs. In other cases, when there are not enough beliefs orienting toward the course of action in at least two belief types, no CO cluster will be formed. The process will discontinue, especially if there are no personally relevant beliefs in the cluster. Similarly, when there is a paucity of beliefs of a certain type, an incomplete CO cluster will be formed which cannot be acted upon but may be discarded or stored for possible use in another situation. Another resulting possibility is the occurrence of conflict (‘intent conflict’) when two CO clusters and consequently two behavioral intents are formed. Another intriguing possibility consists in the formation of an inoperable cluster due to the inclusion of ‘as if’ beliefs in one or more belief types. If, for example, the ‘as if’ beliefs come to replace other beliefs in the beliefs about self and general beliefs, we may get a CO cluster orienting toward virtual action such as daydreaming.
The fourth stage is initiated by the formation of the behavioral intent, and is focused on the question “how will I do it?” The answer is in the form of a behavioral program, namely, a hierarchically structured sequence of instructions governing the performance of the act, including both the more general strategy (‘program scheme’) as well as the more specific tactics (‘operational program’). Previously formed habits are major components embedded in the programs. An important role is attributed to the behavioral intent at this phase. It guides the selection, retrieval, modification and sometimes construction of behavioral programs, and serves as a frame of reference for supervising program execution and evaluating outcomes.
Different programs are involved in executing an overt molar act, a cognitive act, an emotional response, a daydreamed act, conflict resolution, etc. There appear to be four basic kinds of programs: (a) Innately determined programs, which are involved primarily in the output on the submolar level, e.g., those controlling reflexes or classically conditioned responses; (b) Programs that are partly innate and partly acquired by learning, e.g., those controlling instincts (as described by LORENZ 1965, or TINBERGEN 1951), behaviors corresponding to defense mechanisms, and language behavior; (c) Programs acquired through learning, e.g., those controlling culturally shaped behaviors (running elections, etc.), or personal habits (e.g., forms of relaxing, making friends, arranging one’s cupboard); and (d) Programs constructed by the individual ad hoc, in line with situational requirements, often using existing program schemes, including plans for plan construction.
Implementing a behavioral intent by a program requires selecting and retrieving a program, and often adapting it to prevailing circumstances. A ‘program conflict’ may occur between two equally adequate programs or between one that is about to be enacted while another is still in operation.
The retrieval, adaptation and execution of the program, as well as determination of the end-point in view of the behavioral intent also require cognitive control.
Major Advantages of the CO Theory
The major cited advantages will be considered from the perspective of the human level, the level for which the theory was originally developed and in which it was primarily applied. The advantages are partly applied and partly theoretical.
The major advantage from the applied viewpoint is that the CO theory enables predictions of behaviors and changes in behavior by using systematic procedures.
Evolution and Cognition ❘ 86 ❘ 2001, Vol. 7, No. 1
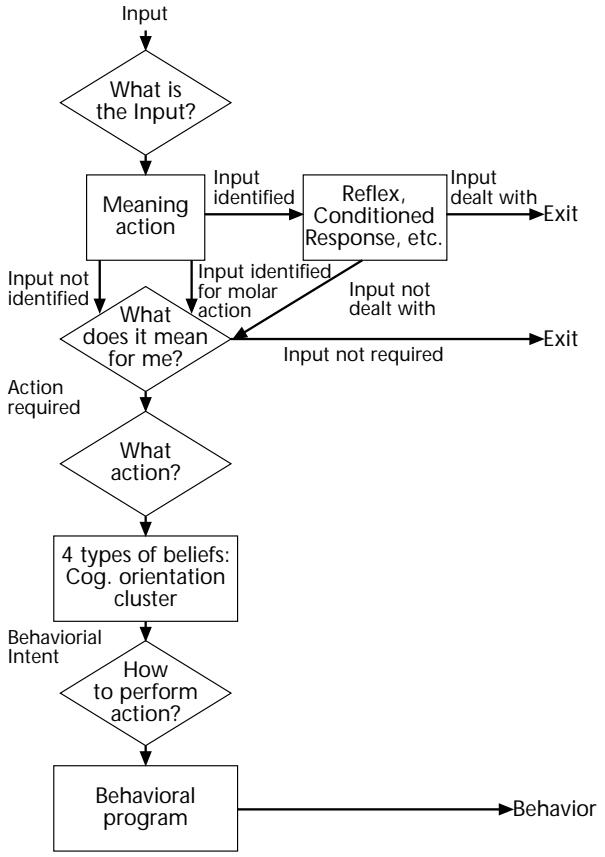
Figure 3: A Schematic Flow-Chart of the CO Model
A large body of research demonstrates the predictive power of the CO theory in a variety of domains, e.g., coming on time, reactions to success and failure, curiosity, achievement, studying mathematics, planning, assertiveness, conformity, cheating, overeating, breast-feeding, cessation of smoking, self-disclosure, rigidity, defensive responses, undergoing tests for the early detection of breast cancer, sexual responses, compliance in diabetes patients etc. All studies refer to actual observed behaviors. The participants were adults, adolescents, children, retarded individuals, schizophrenics, individuals with different physical disorders, etc. (KREITLER/CHAITCHIK/KRE-ITLER/WEISSLER 1994; KREITLER/CHEMERINSKI 1988; KREITLER/KREITLER 1976, 1982, 1988, 1991a, 1993, 1994a, 1994b; KREITLER/NUSSBAUM 1998; KREITLER/ SCHWARTZ/KREITLER 1987; KREITLER/SHAHAR 1976; LO-BEL 1982, NURYMBERG/KREITLER/WEISSLER 1996; TIP-TON/RIEBSAME 1987; WESTHOFF/HALBACH-SUAREZ 1989).
All studies confirmed the hypothesis that behavior would occur if it was supported by at least three
| Themes | Beliefs about Self |
Beliefs about Norms |
Beliefs about Goals |
General Beliefs |
|---|---|---|---|---|
| 1) | ||||
| 2) | ||||
| 3) | ||||
| : | ||||
| n) |
Figure 4: The Predictive Matrix of Beliefs.
belief types and a behavioral program was available. The success of the predictions is based on applying the special standardized procedure developed in the framework of the CO theory (KREITLER/KREITLER 1982; in press). The procedure consists in assessing the motivational disposition for the behavior (viz. behavioral intent) by means of a CO questionnaire and examining the availability of a behavioral program for implementing the intent. A CO questionnaire assesses the degree to which the participant agrees to relevant beliefs orienting toward the behavior in question or rejects those that do not orient toward it. The beliefs differ in contents and form. In contents they refer to themes which represent meanings underlying the behavior in question (called ‘themes’). In form they refer to the four types of beliefs, namely, beliefs about goals, beliefs about rules and standards (or norms), beliefs about the self, and beliefs about others and reality (called general beliefs). Thus, a CO questionnaire mirrors the prediction matrix (see Figure 4). It usually consists of four parts presented together in random order, each representing one of the four types of beliefs and containing in random order beliefs referring to the different themes. The participant is requested to check on a 4-point scale the degree to which each belief seems to him/her true (or correct).
The themes of the CO questionnaire are identified by means of a standard procedure applied to pretest participants who are known to manifest the behavior in question or not. The procedure consists in interviewing the participants about the meanings of the key terms and then in turn sequentially about their responses concerning the meanings (see Figure 5 for a schematic representation of the procedure). Repeating the questions about meanings leads to deeper-layer meanings, out of which those that recur in at least 50% of the interviewees are selected for the
Evolution and Cognition ❘ 87 ❘ 2001, Vol. 7, No. 1
| Interpersonally-shared meaning of behavior = key words |
|||
|---|---|---|---|
| Level 1: _ |
Personal meanings of key-words | ||
| Level 2: _ |
Personal meanings of responses on level 1 | ||
| Level 3: | Personal meanings of responses on level 2 |
Figure 5: The Process of Identifying Themes by means of Meaning Generation.
final questionnaire. Examples of themes for success in dieting are acceptance of limitations, and viewing the world as a safe place. It is important to emphasize that the beliefs in the questionnaire do not refer in any way to the behavior in question but only to the themes which represent the underlying meanings. Identifying the themes is followed by constructing a CO questionnaire, which has to be examined for its psychometric properties, including reliability and validity before it is ready for application. Although for every kind of behavior it is necessary to construct a particular CO questionnaire, a single CO questionnaire may predict a broad range of relevant behaviors, e.g., the CO questionnaire of curiosity predicted 14 different curiosity behaviors (KREITLER/KREITLER 1994b).
The availability of the behavioral program is established by means of questionnaires, observation, information from others or role-playing.
The advantages of the prediction procedure generated by the CO theory are the following: (a) it has provided a great number of significant predictions of actual behaviors; (b) the predicted behaviors were of many different kinds and from different domains; (c) the participants whose behaviors were predicted were of different ages and intelligence levels, even cognitively challenged individuals; (d) applying the procedure does not require any special ‘mind-set’, preparation, intention, or even average intelligence on the part of the participants; (e) the procedure does not enable the participants to tailor their responses so that it would or would not correspond to their behaviors because it is impossible to unravel from the statements in the CO questionnaire the kind of involved behavior; (f) the procedure may be applied in regard to any behavior whatsoever provided that it can be assessed; (g) applying the prediction procedure is straightforward and does not require the involvement of further criteria or constructs or assumptions or the creation of particular conditions in regard to any behavior.
The CO theory has also enabled successful modifications of behavior, such as rigidity, impulsivity, curiosity and eating disorders (BACHAR/LATZER/ KREITLER/BERRY 1999; KREITLER/KREITLER 1988, 1990C, 1994B; ZAKAY/BAR-EL/KREITLER 1984). The essential components of the procedure consist in mobilizing sufficient support for a specific course of action by evoking in the participant beliefs orienting toward this course of action. The major conditions on which success of the intervention depend are that the evoked beliefs (a) refer to the themes (viz. underlying meanings of the action), (b) refer to all four types of beliefs (viz., goals, norms, self and general), and (c) that they have originated in the participant by a process of meaning generation. The procedure for intervention or change of behavior has been standardized and can be applied in individual or group sessions (KREITLER/KREITLER 1990c). Again, there are no limitations on the types of behaviors or of participants in regard to which it can be applied.
The major theoretical advantages of the CO theory are that it shows precisely what is the role of cognition in regard to motivation and behavior. The assumption that attitudes, values, informations and opinions are important in regard to behavior is deeply rooted in Western culture and underlies education, persuasion and dissemination of information through the media and otherwise. However, a large body of studies in the last century showed that attitudes were not related to behavior (e.g., COREY 1937; LAPIERE 1934). This evidence posed a serious problem for behavioral scientists (KREITLER 2002). The CO theory provided a plausible and operational account of the role of cognition in regard to motivation and behavior. Thereby, the CO theory also clarified the role of several additional basic concepts in regard to motivation and behavior, mainly the self, values and norms, and attributions, causal and others.
Evolutionary Aspects of the CO Theory
The evolutionary aspects of the CO theory will be discussed in two complementary forms. The first will be in terms of general developmental trends, the second—in terms of the stages intervening between input and output in line with the CO theory (see section “The CO theory: How does it function? Major theoretical stages”). In both cases the discussion will be based partly on extrapolations from observations on ontogenetic development, and partly on information about animal behavior. However it may be, the purpose is merely to trace
Evolution and Cognition ❘ 88 ❘ 2001, Vol. 7, No. 1
the blueprint of evolutionary aspects involved in the cognitive guidance of behavior.
General developmental trends. The first and possibly the most important trend is the differentiation of action and motivation. This differentiation does not seem to exist at the lowest evolutionary levels. On these levels we find actions, which may be prompted by inner or outer states that are not differentiated from the action and cannot be identified as such. As stated most precisely by WIMMER (1995, p43), “information about [homeostatic] disturbance and motoric pattern are combined in a unit which becomes segregated at higher levels of organization, e.g., taxis behavior”. The differentiation occurs when we first obtain some kind of representation of the inner or outer stimulus. This constitutes the basis for the development of motivation in later phases, as distinct from action. The two arms—of motivation and of action—undergo different transformations and development in the course of evolution. The development of motivation depends largely on meaning, whereas the development of action depends largely on structures.
Accordingly, the second basic trend is the evolution of meaning. This evolution takes different forms, some of which overlap, whereas others occur in a sequence. The development is partly structural and partly dynamic. The structural development consists basically in shaping the major outlines of the meaning system. This entails differentiating the contents of the meaning values into subsets which we call meaning dimensions, differentiating the relations of the contents to the referents in terms of directness, generality, positivity and the nature of the relation (propositional, normative, desired), and moving from the nonverbal actional means of expression to the use of diversified nonverbal and verbal means of expressing meaning values and meaning units. On higher levels, the whole system splits into two major branches of the interpersonallyshared meaning and the subjective–personal meaning, which may intertwine and interact to different degrees. The more dynamic development consists in moving from meaning values to meaning units (each of which includes a meaning value centered around a referent), from meaning action to meaning generation, and from surface or label meaning to elaboration of the deeper underlying meanings. The latter development brings aboutan increasing involvement of personal–subjective meaning in addition to the interpersonally-shared one which dominates the surface-label meaning.
A special line of development is taken by meaning units. It will be mentioned because it is of particular importance for the cognitive guidance of motivation. This line leads from meaning units to beliefs. Both meaning units and beliefs share the basic structure of a referent related to a meaning value. However, they differ in function. Meaning unit is a concept from the domain of meaning and its function is the expression or communication of meaning per se. In contrast, belief is a concept from the domain of action or behavior, and its function is to express or communicate some cognitive contents—e.g., a statement, a view, an attitude, an opinion, an information, a value, an expectation etc.—with behavioral implications.
The primary meaning unit is a belief which states a fact, namely, the relation of the meaning value to the referent is factual, existential. Hence, it could be either a belief about self or a general belief. It is difficult to decide which one was the first. But in view of the primary focusing on oneself it seems likely that the belief about self preceded the general belief. Be it as it may, each of these primary units split to form by differentiation two distinct varieties. Thus, the belief about self differentiated into a belief about goals and a belief about self, both of which share the ‘I’ as a referent and differ only in the kind of relation between the meaning value and the referent (viz. desired vs. factual, respectively). Similarly, the general belief differentiated into a belief about norms and a general belief, both of which share the non–I as a referent and differ only in the kind of relation between the meaning value and the referent (viz. normative/desirable vs. factual, respectively). As a result, four types of beliefs emerged.
Complementing the evolution of meaning is the third trend, which refers to the evolution of action. The major characteristics of evolution in this sphere have been extensively studied. One characteristic consists in increased variety of action units. On the lower evolutionary levels we find genetically and innately determined units of action that are rigidly enacted in a specific manner. New units are added to the repertory which modify the innately given actional units or combine with them in different ways and thus form, for example, conditioned responses and habits. Further, the units of action which initially may be bound to specific inputs or situations become gradually free of such constraints, and may be used in a variety of contexts, or for the attainment of purposes completely divorced from those with which they may have been related when acquired. Another characteristic development consists in in-
Evolution and Cognition ❘ 89 ❘ 2001, Vol. 7, No. 1
creased complexity of organization of the actions. For example, several action units may combine into larger units by forming a sequence or chain of units or a hierarchically structured ‘family of habits’, wherein one or more action units may be replaced by other units when the former do not attain the goal.
The increased organizational complexity enables also increased flexibility in form, structure and enactment, which is manifested, for example, by selecting action units in line with situational characteristics and adapting the enactment of the action units to particular contextual conditions in which the action takes place. Increased flexible adaptability may reach a level where its implementation cannot depend any more on tuning-in with the situational constraints as one goes along, following the trial– and–error method. The next major evolutionary step in this respect consists in the emergence of the behavioral program which completely changed the scene of action by introducing from above an organizational principle that takes control over action from “the top, organizing the enactment in terms of an overall scheme”. This change may have occurred at the level of worms. The program starts to play a role in the selection of the appropriate action units for the behavior and in the way habits are embedded within it (GALLISTEL 1980; MILLER/GALANTER/PRIBRAM 1960).
Within the sphere of behavioral programs a further evolution takes place whereby learning gets an increased role and contribution. The primary programs are innate ones, such as reflexes. These programs become slowly more flexible and adaptable. For example, conditioning brings about the binding of these primary programs to new signals so that they come to be evoked by learned stimuli in the context of new situations. These and other processes of learning invade, as it were, the innate schemes, enlarging and transforming them in different ways. Moreover, the innate programs are gradually amplified by newly evolving programs that are increasingly based on learning. Thus, the next stage consists of programs that are partly innate and partly learned, such as instincts, behaviors corresponding to defense mechanisms, language and social behaviors. The components due to learning become gradually more salient and push the innately-given components into the background so that they become barely perceptible. For example, in an instinct the learning-based ‘appetitive behavior’ is distinctly differentiated from the end-stage behavior which represents the more innate components. However, in a defense mechanism behavior such as displacement, the innate element is represented not in the actual manifest behavior but in its underlying cause or goal. In different social behaviors, such as courting, threatening or impressing others the innate element is even more covert (HINDE 1970, pp370–382; MORRIS 1969).
The next evolutionary stage is characterized by the emergence of behavioral programs that are purely learned, such as, driving a car or using the internet. It is likely that even here there are some barely perceptible but identifiable innate elements that have stealthily invaded the scheme (e.g., pushing buttons, turning wheels). The next evolutionary stage consists in giving up ready-made programs and relying increasingly on ad-hoc programs newly created in line with the present circumstances. Thus, program retrieval is gradually replaced by planning. A series of studies on the development of planning showed that the major developments occur in regard to information gathering (focusing on items of information necessary for proceeding smoothly with the action), information organization (for example, in terms of if–then considerations, or defining domains of information), and plan structuring (chronological ordering, number of steps, major and alternate plans, etc.). The development of planning and acquisition of planning skills are promoted by motivation and increased awareness of needs (KRE-ITLER/KREITLER 1987a–c).
The advantage of the ad-hoc programs is their perfect fit to the situation, their disadvantage—that they have to be created under the pressure of ongoing behavior. The task is however made easier in view of the more advanced development of programs for the formation of programs. These programs make use of elements accumulated in the course of previous development, from the bottom, as it were, but as soon as they emerge they start controlling the processes from the top, as it were. However, in this field too, the more developed manifestations do not completely replace the lower-level products, which may still be adequate for handling specific situations more adequately than higher-level behaviors (see the principle of “preservation of the fittest” in “The CO Theory: General Characteristics”).
The described development of programs emphasized their increased shaping by learning. The development of programs may also be examined from the complementary viewpoint of the changes they undergo. First, it is evident that behavioral programs increase in number, so that the organism has programs adequate for an increasing number of differ-
Evolution and Cognition ❘ 90 ❘ 2001, Vol. 7, No. 1
ent situations, and also more programs for the same or similar situations from which one can select the adequate one. Secondly, behavioral programs grow in complexity in the sense that they get more hierarchically organized with an increasing number of levels. This involves not only an increasing number of units encompassed within the program but also consideration of special conditions, alternatives and ‘if then’ eventualities. With time, following differentiation processes in complex systems (RIEDL 1997, p152), a differentiation occurs between the levels that deal with the overall strategy of action (viz. ‘operational program’) and the levels that deal mainly with the operational details involved in the specifications of the enactment, mostly in terms of physiological processes (viz. ‘program scheme’). The increased complexity of organization enables activating not only the program as a whole but parts of programs, or shaping new programs by integrating parts from other programs. At a higher level of development, behavioral programs themselves undergo further evolution in that they are organized, for example in terms of similarity or relevance for specific situations. Eventually this leads to the third important development which consists in forming principles for the construction of programs. This frees performance to a certain extent from reliance on ready-made programs and introduces the option of planning, with all the freedom and control it involves.
Specific developmental trends. The present section is devoted to highlighting diverse evolutionary trends affecting the phases defined by the CO model as intervening between input and output. Since the CO model is a cognitive model of motivation, tracing the evolutionary trends involved in the phases of the model would highlight more specifically than before the evolutionary development of the cognitive guidance of motivation.
In the first phase, the primary actions would be activities dependent completely on internal stimuli, if indeed one may consider the internal perturbations of disturbed homeostasis as stimuli. In the best case, they are equivalents or precursors of stimuli. The differentiation between the motivation and action starts at this phase. WIMMER (1995) provides a most original and detailed reconstruction of the differentiation in evolutionary perspective. At the lowest level we start with the primary action unit, whose activity is regulated by internal stimuli. These internal stimuli (the earliest precursor of emotion), reflect both the inner state of the organism (homeostasis or
‘disturbance’) and the outer state (they are the ‘perception’ of the environment mediated by the organism’s homeostasis, p43). Moreover, they contain the roots of motivation in the form of ‘expectation’ of information and of response, in the form of rigid modes of reaction (RIEDL 1980). The evolutionary development shows that originally the locomotory activity of kinesis, for example, in taxis behaviors, is rigid, automatically regulated and appears to be undirected. At higher levels of taxis behaviors it is replaced by a type of action which shows the earmarks of being directly determined by the impinging stimulus (LORENZ 1981), reflecting the impact of the organism’s internal state. This is probably the earliest precursor of goal-directedness. Notably, it corresponds to ‘intentionality’, that has been identified as a basic characteristic of behavior, at least from the level of organisms evidencing directed movements or taxes (DIAMOND 1991; FREEMAN 1999, p41). The manifestation of intentionality marks the emergence of motivation as distinct from the action per se. WIMMER (1995, p43) cites in this context the example of the behavior of the turbellarian worms whose movement toward a feeding site depends on their internal state: the hungry worm will move toward the site, whereas a satiated worm will not. Hence, WIMMER concludes that it is the differentiation of the internal state (e.g., hungry, satiated) which leads to the development of more different types of perceptions (sensitivity to different constellations of stimuli) and of motoric reactions (approach or avoidance reactions). With the development of perception, it becomes possible for external stimuli to initiate action. In terms of the CO theory, it is the motivational disposition which drives development of the grasp of reality and of the performance component.
Further evolutionary trends that may be relevant in regard to the first phase of the CO theory depend on the development of meaning elaboration and feedback evaluation. At the lowest level, the input which initiates action has meaning which is minimal (it consists perhaps of only one meaning value) and is couched in actional terms. Hence, we call it ‘meaning action’. Its manifestation is precisely in evoking action, say, a reflex. It consists in identifying the input as a signal for a specific action, namely, in evoking action and it has no independent existence outside action. This applies to stimuli that evoke reflexes or primitive defensive reactions. ‘Releasers’ (or sign stimuli) are at a slightly higher evolutionary level insofar as they act as triggers for instincts, which are chains of actions manifesting some degree
Evolution and Cognition ❘ 91 ❘ 2001, Vol. 7, No. 1
of plasticity. At higher evolutionary levels we may find meanings that are not identical with a specific action, for example, a meaning that is evaluated as irrelevant and hence is dismissed without evoking any response, or a meaning that is evaluated as requiring a more complex type of behavior and hence is transferred to the next phase.
Similarly, also feedback evaluation is subject to development. Basically, feedback evaluation depends on applying cognitive processes and meaning. Thus, evaluating the outcome of action requires matching the outcome with the intent, adjusting both so that they can be compared, judging the degree of similarity, and evaluating the result also in terms of the current situation and its potentialities. This description fits higher evolutionary levels. On lower levels the precursor of feedback would consist simply in recurrent enactment of the action if the disturbance exists, regardless of whether the action had been enacted before and what the goal may have been. The action is simply repeated until the goal is attained or a ‘stop circumstance’ sets in (e.g., the organism is distracted, the organism stops because of fatigue, the action is inhibited by an internal inhibiting mechanism). In this context it is relevant to cite the observations about infants 5–7 months old trying again and again without success to retrieve a toy they had and which was taken away from them (DI-AMOND 1991). Hence, at low evolutionary levels, it may happen that an action does not attain the goal in any sense, yet it is not followed by another action. At a later evolutionary level, feedback already in the case of instincts is based on a rigid type of matching the actual activity or outcome with the original ‘expectation’ or ‘optimal’ pattern. This geneticallybased yardstick (WIMMER 1995, p45) or ‘inborn teacher’ (LORENZ 1981, p299) is amplified by more flexible and extended forms of feedback, based on learned programs stored in memory. One type consists in evaluating the correspondence between actual action and stored pattern (KLIX 1980, p102), another in estimating the accuracy of reaching the goal (ibid, p104). WIMMER (1995) emphasizes that the latter is anchored in the brain centers of reward and punishment, so that it uses elementary emotional reactions as criteria for success and failure of actions. The development of these feedback mechanisms has helped to dissociate behaviors from specific stimuli and has promoted the occurrence of behaviors in contexts other than those in which they have been acquired (e.g., conditioned actions) (ibid, pp45–46). With further developments the feedback mechanisms become more flexible and expanded, consisting in examining the outcome situation more extensively and comparing it in several senses with the original goal(s). At this level, the feedback operation already includes the vestiges of preparing the next action designed to handle the original goal in view of the experienced failure of the previous action.
In the first phase of the CO theory there are at least two occasions for feedback: after the execution of the non-conditioned or conditioned response, and after evocation of the orienting reflex. The previous observations about the evolution of feedback indicate that it requires higher evolutionary levels to diagnose failure in these actions, and in case of failure, it requires even higher levels to transfer the handling of the situation to more advanced phases (viz. to molar action).
In line with the CO theory, the second phase is focused on a more extended elaboration of the meaning of the input and the situation in which it is embedded. This phase is designed to examine, on the bases of the meanings which emerge, whether action is at all indicated. This phase bears evidence for an advanced evolutionary level. In addition to an extended meaning system, it presupposes the ability not to act. Studies with infants suggest that this ability is based on a more basic ability, which is not to react in line with innate and conditioned prepotent response tendencies. The latter gives one the possibility of reacting in line with internal tendencies. Extending the potential of inhibiting actions to planned, considered and possible actions, largely widens the scope of intentionality and control. Both abilities of inhibiting action are probably not possible without the frontal cortex (DIAMOND 1991).
There seem to be two major evolutionary trends characteristic of the second phase. The first and most obvious one concerns meaning elaboration. The broader and more comprehensive the meaning system becomes, the more extensive gets the meaning elaboration in this context, involving more aspects of meaning and both types of meaning—the interpersonally-shared and the personal, in the form of meanings as well as beliefs. Correspondingly, the examination of whether the individual is involved in the situation is based on an ever-broader basis. The second aspect that undergoes evolution is the criteria for acting or not acting. If on more primary levels the decision follows criteria, such as direct contribution to survival, evolution brings into play further criteria, such as potential for promoting the attainment of some personal goal, the possibility to defend one’s values or to enhance one’s sense of self-identity, etc.
Evolution and Cognition ❘ 92 ❘ 2001, Vol. 7, No. 1
The third phase is focused on determining the directionality of behavior. One important evolutionary trend concerns the emergence of underlying meanings. Their range and depth increase so that they come to represent the matrix of meanings out of which the behavioral intent can be shaped. This however raises a dilemma. On the one hand, it is desirable for shaping the behavioral directionality to evolve a broad and deep set of underlying meanings. Yet, on the other hand, awareness of this complete set would render action difficult insofar as it constitutes a heavy memory load and uses large mental resources of consciousness. The solution to the dilemma could be to develop the whole matrix but to keep it unconscious. Possibly, the demands of smooth action and preservation of mental resources may have promoted development of the unconsciousness.
Other interesting lines of development concern the formation of the CO cluster. Most important of all is the issue of the number of belief types that are necessary for creating a CO cluster. At the lowest level, as a rule only one belief type dominates the scene, most probably goals (= representing approach or avoidance, “I want” or “I don’t want”). The next belief type to make its contribution to behavioral directionality is rules and norms (= representing constraints of right and wrong, allowed or forbidden and later also of utility, desirability etc.). Each could dominate the scene in itself. An important development occurs however when beliefs about goals and beliefs about rules and norms do not point in the same direction, or when they clash. A study with children showed that it is only under these conditions that the other belief types general beliefs and self beliefs enter the scene and begin to make a difference in predicting the child’s behavior (MAROM 1978).
There are other pairs of belief types that could form inconsistencies of consequence for the development of the CO cluster. One is the inconsistency between beliefs about goals and general beliefs, which occurs when prevailing circumstances do not support the attainment of a goal. This may have been the case for the apes studied by DELGADO (1974) whose goal to express aggression was not matched by the information that the target or would-be victim is a strong ape who would not easily succumb. Another example is the inconsistency between beliefs about norms and beliefs about self, which occurs for example when one becomes aware of not living up to the requirements. Again, there is the gap between beliefs about norms and general beliefs that
may occur when reality does not live up to normative requirements. A last example concerns the gap between beliefs about self and general beliefs, which occurs when informations about oneself and others are inconsistent. In each of these cases the drive for solution is based on bringing into consideration additional information, in the form of further types of beliefs. Hence, the drive behind the formation of a CO cluster representing the overall directionality implied by four types of beliefs is the inconsistencies in directionality between any two pairs of belief types. Studies showed that when individuals function under difficult conditions, such as the impact of drugs or medication, drunkenness, fatigue, or intense emotions, they tend to form CO clusters based on fewer than all four types of beliefs. The inconsistencies between belief types serve to mobilize for the CO cluster more beliefs. This enables forming a CO cluster if there are enough beliefs supporting the course of action rather than if all beliefs or all belief types support it.
The increase in the number of belief types on which the CO cluster is based is probably attended also by other developments in the formation of the CO clusters. We will mention in particular four that are of special importance. The first concerns the anchor of the CO cluster. Originally, the anchor must have been a goal belief. In the course of development, other belief types could assume this role, in line with the characteristics of the behavior considered for enactment. For example, general beliefs when the behavior is conformity, norm beliefs when the behavior is helping others, beliefs about self when the behavior is assertiveness. Secondly, also the salience of the belief type in the formation of the CO cluster developed from a fixed order to an order that varies in line with the behavior and circumstances, and later possibly also personality. Thirdly, the range of implications of the beliefs that exerts an impact on the processes of belief interactions and clustering undergoes change. The primary and most important kind of implication is orientativeness, namely, the degree to which the belief supports the behavior. Development sets in operation further types of implications (e.g., morality, utility, emotional impact), which though initially they play a secondary role, they increasingly contribute to the formation of the CO cluster and to shaping the directionality of behavior. Finally, development occurs also in regard to the amount of consistency among the orientativeness of the beliefs required for the formation of a CO cluster and the consequent emergence of the behavioral intent. On the lowest
Evolution and Cognition ❘ 93 ❘ 2001, Vol. 7, No. 1
developmental level of the CO cluster the requirement is for perfect or almost perfect consistency in the orientativeness of the beliefs. All relevant beliefs must support the behavior; otherwise no CO cluster is formed. With evolution the standards of requirements are loosened, until the stage is reached when consistency is required only within the CO cluster but not among all beliefs relevant for the behavior. Thus, the primary question “Do all beliefs support the behavior?” is replaced by the question “Is there sufficient support for the behavior?” This development enables the accumulation or generation of beliefs of all kinds, without prior consideration of the behaviors which they could support or not. Subsequently, further beliefs are generated or acquired without regard for their actual or potential contradiction with the already existing beliefs. Removal of the constraints on belief accumulation enables a great enrichment of cognitive contents. The impact on cognitive development is remarkable.
Furthermore, it can be expected that also the criterion for ‘sufficient support’ becomes more flexible with time so that assessing sufficiency is based not only on the number of involved beliefs but also on additional aspects, such as the situational context, kind of behavior, and meaningfulness of the beliefs
The fourth phase is focused on behavior performance. The major evolutionary developments refer to the activation, adaptation and enactment of the behavioral programs.
Activation involves identifying and evoking the adequate program. On the lower evolutionary levels, each behavioral program is related to a specific stimulus or range of stimuli. Under these conditions, identifying the adequate program consists in retrieving it as a response when the proper stimulus appears. Identifying and retrieving are not yet differentiated. When behavioral programs undergo dissociation from the releasing stimuli, retrieval becomes a matter of search in memory for the adequate program. The selection process involves focusing on a pattern of behavior that is marked as ‘adequate’ and searching in memory for a pattern that matches the constructed concept to a satisfying or ‘satisficing’ degree. The ‘adequate’ program is initially searched from among those stored in memory, and in later evolutionary levels may be shaped or constructed ad hoc.
Adaptation of programs involves an interaction between the program and the available circumstances that results in transforming the program so that it increasingly matches the circumstances. Development brings about an ever-increasing adequacy in the fit between the behavioral program and the context in which the behavior takes place. Accordingly, adaptation consists in a gradual departure from rigid action units or sequences in favor of flexible, changing, and variable behaviors. The difference becomes most apparent when one compares the relative flexibility in the operation of instincts, especially in the appetitive stage, with the rigidity in the operation of reflexes. It is as if the stored behavioral program becomes a paradigm for a set of variations that assume unique shapes when set in operation in reality.
Enactment of programs involves an interaction between the program and the emerging reality contingent upon setting it in operation. Initially, enactment is examined only in terms of the end-point (i.e., has the action been terminated or not). Later developments introduce control over intermediate and primary stages of enactment. Control consists first in determining whether the action takes place or not, but it evolves into a more complex operation which includes also evaluation of the operation itself and its outcomes and the possibility of modifying it, if necessary. Thus, control assumes gradually an autonomous status from the performance itself. Thereby it becomes increasingly more cognitive and enables the introduction of additional criteria for the implementation of control. On the lower evolutionary levels the main criteria for evaluating ongoing performance is the occurrence of the activity itself (namely, whether the action occurs and runs to its end), implemented by utility (the pragmatic evaluation in terms of whether the straightforward goal was attained). Evolution enables the introduction of further criteria, mainly whether the action and/or the goal attainment are emotionally satisfying, whether they conform to social desirability, whether they are useful also in promoting the attainment of other goals, whether they conform to standards of reasonableness or rationality, whether they were sufficiently economical in the use of resources (such as, time, money, energy), whether they satisfy aesthetic requirements, etc. Notably, the application of considerations of this kind for evaluating the performance and its outcomes serve to enhance the integration of behavior into the system of personality.
Conclusions
One major conclusion of this paper is that the cognitive guidance of behavior is an evolutionary product, reflecting multiple developmental trends. Major among these are the differentiation between
Evolution and Cognition ❘ 94 ❘ 2001, Vol. 7, No. 1
motivation and action, the development of meaning, and the development of action. In the course of evolution, motivation unfolds in a manner that enables its increasing suffusion and consequent shaping by cognitive contents and processes anchored in the components and dynamics of meaning. Meaning assignment processes are, however, subjected to the overall motivational framework that determines the purpose, manner and products of the meaning elaboration. This means that cognitive elaboration in the context of motivation differs in contents and processes from cognitive elaboration in other contexts. Thus, in contrast to ‘pure’ problem solving and other contexts in which cognition is operative, meaning elaboration in the motivational framework capitalizes on the following: a specific balancing of interpersonally-
shared and personal–subjective meaning; the use of meaning action; treating meaning units as beliefs; activating four types of beliefs; unraveling underlying meanings of the inputs to a specific level of depth; focusing on
Author’s address
Shulamith Kreitler, Department of Psychology, Tel-Aviv University, Tel-Aviv 69978, Israel, Tel: +972-3-5227185, Fax: +972-3- 5225371. E-mail: krit@netvision.net.il
orientativeness of beliefs as a particular aspect of the meaning of beliefs; and clustering of beliefs into a CO cluster expressible in terms of a goal-directed vectorial disposition.
In the course of evolution, action develops in the following ways: organizing action units; freeing them from original inputs or inputs related to their acquisition; increasing complexity of organization; development of behavioral programs, amplifying the innate programs by partially-learned and fullylearned programs; amplifying and replacing stored programs by planning, which produces ad-hoc plans, some of which may be stored for future use; cognitive control of the enactment of programs, including the evaluation of action outcomes.
In view of the above, it is justified to conclude that the cognitive guidance of behavior underscores and
renders explicit an important manifestation of the contribution of cognition to behavior and to survival. Thus, it highlights cognition’s unique role in terms of evolution, as it is shaped by evolution, for evolution.
References
- Ajzen, I. (1985) From intentions to actions: A theory of planned behavior. In: Kuhl, I./Beckman, J. (eds) Action Control: From Cognition to Behavior. Springer: Berlin, pp. 11–40.
- Bachar, E./Latzer, Y./Kreitler, S./Berry, E. M. (1999) Empirical comparison of two psychological therapies: Self psychology and cognitive orientation in treatment of anorexia and bulimia. Journal of Psychotherapy Practice and Research 8: 115–128.
- Becker, M. H. (1974) The health belief model and personal behavior. Health Education Monographs 2: 324–508.
- Campbell, D. T. (1974) Evolutionary epistemology. In: Schlipp, P. A. (ed) The Philosophy of Karl Popper. Open Court: LaSalle IL, pp. 413–463.
- Corey, S. M. (1937) Professed attitudes and actual behavior. Journal of Educational Psychology 28: 271–280.
- Cassirer, E. (1953) An Essay on Man: An Introduction to a Philosophy of Human Culture. Doubleday, Anchor Books: Garden City NY.
- DeKay, W. T./Shackelford, T. K. (2000) Toward an evolutionary approach to social cognition. Evolution and Cognition 6: 185–195.
- Delgado, J. M. R. (1971) Physical Control of the Mind: Toward a Psycho-Civilized Society. Harper: New York.
- Diamond, A. (1991) Neuropsychological insights into the meaning of object concept development. In : Carey, S./ Gelman, R. (eds) The Epigenesis of Mind: Essays on Biolo-
gy and Cognition. Erlbaum: Hillsdale NJ, pp. 67–110.
- Freeman, W. J. (1999) How Brains Make up their Minds. Phoenix, Orion Books: London.
- Gallistel, C. R. (1980) The Organization of Action: A New Synthesis. Erlbaum: Hillsdale NJ.
- Gallistel, C. R. (1990) The Organization of Learning. A Bradford Book, MIT Press: Cambridge MA.
- Hinde, R. A. (1970) Animal Behavior: A Synthesis of Ethology and Comparative Psychology. 2nd edition. McGraw-Hill: New York.
- Klix, F. (1980) Erwachendes Denken. Eine Entwicklungsgeschichte der menschlichen Intelligenz. VEB Deutscher Verlag der Wissenschaften: Berlin.
- Ková , L. (2000) Fundamental principles of cognitive biology. Evolution and Cognition 6: 51–69. cˇ
- Kreitler, S. (1965) Symbolschöpfung und Symbolerfassung: Eine experimentalpsychologische Studie [The Creation and Perception of Symbols: An Experimental Psychological Study]. Reinhardt: Basel, München.
- Kreitler, S. (1999) Consciousness and meaning. In: Singer, J. A./Salovey, P. (eds) At Play in the Fields of Consciousness. Erlbaum: Mahwah NJ, pp. 175–206.
- Kreitler, S. (2002) The cognitive guidance of behavior. In: Jost, E. et al. (eds), The Yin and Yang of Social Cognition: Perspectives on the Social Psychology of Thought Systems. Erlbaum: Mahwah NJ.
- Kreitler, S./Chaitchik, S./Kreitler, H./Weissler, K. (1994) Who will attend tests for the early detection of breast cancer? Psychology and Health 9: 463–483.
Evolution and Cognition ❘ 95 ❘ 2001, Vol. 7, No. 1
Kreitler, S./Chemerinski, A. (1988) The cognitive orientation of obesity. International Journal of Obesity 12: 471–483.
- Kreitler, H./Kreitler, S. (1965) Die weltanschauliche Orientierung der Schizophrenen [The Cognitive Orientation of Schizophrenics]. Reinhardt: München, Basel.
- Kreitler, H./Kreitler, S. (1972) The model of cognitive orientation: Towards a theory of human behavior. British Journal of Psychology 63: 9–30.
- Kreitler, H./Kreitler, S. (1976) Cognitive Orientation and Behavior. Springer: New York.
- Kreitler, H./Kreitler, S. (1982) The theory of cognitive orientation: Widening the scope of behavior prediction. In: Maher, B. A./Maher, W. A. (eds) Progress in Experimental Personality Research: Vol. 11. Normal Personality Processes. Academic Press: New York, pp. 101–169.
- Kreitler, S./Kreitler, H. (1985) The psychosemantic foundations of comprehension. Theoretical Linguistics 12: 185– 195.
- Kreitler, S./Kreitler, H. (1986) Individuality in planning: Meaning patterns of planning styles. International Journal of Psychology 21: 565–587.
- Kreitler, S./Kreitler H. (1987a) The motivational and cognitive determinants of individual planning. Genetic, Social, and General Psychology Monographs 113: 81–107.
- Kreitler, S./Kreitler, H. (1987b) Plans and planning: Their motivational and cognitive antecedents. In: Friedman, S. L./Scholnick, E. K./Cocking, R. R.(eds) Blueprints for Thinking: The Role of Planning in Cognitive Development. Cambridge University Press: New York, pp. 110– 178.
- Kreitler, S./Kreitler, H. (1987c) The development of planning. In: Friedman, S. L./Scholnick, E. K./Cocking, R. R.(eds) Blueprints for Thinking: The Role of Planning in Cognitive Development. Cambridge University Press: New York, pp. 205–272.
- Kreitler, S./Kreitler, H. (1988) The cognitive approach to motivation in retarded individuals. In: Bray, N. W. (ed) International Review of Research in Mental Retardation, Vol. 15. Academic Press: San Diego CA, pp. 81–123.
- Kreitler, S./Kreitler, H. (1989) Horizontal decalage: A problem and its resolution. Cognitive Development 4:89–119.
- Kreitler, H./Kreitler, S. (1990a) The psychosemantic foundations of creativity. In: Gilhooly, K. J./Keane, M./Logie, R./ Erdos, G. (eds) Lines of Thought: Reflections on the Psychology of Thinking, Vol. 2. Wiley: Chichester, pp. 15–28.
- Kreitler, S./Kreitler, H. (1990b) The Cognitive Foundations of Personality Traits. Plenum: New York.
- Kreitler, H./Kreitler, S. (1990c) Cognitive primacy, cognitive behavior guidance and their implications for cognitive therapy. Journal of Cognitive Psychotherapy 4: 155–173.
- Kreitler, S./Kreitler, H. (1991a) Cognitive orientation and physical disease or health. European Journal of Personality 5: 109–129.
- Kreitler, S./Kreitler, H. (1991b) The psychological profile of the health-oriented individual. European Journal of Personality 5: 35–60.
- Kreitler, S./Kreitler, H. (1993) The cognitive determinants of defense mechanisms. In: Hentschel, U./Smith, G./Ehlers, W./Draguns, J. G. (eds) The Concept of Defense mechanisms in contemporary psychology: Theoretical, research and clinical perspectives. Springer-Verlag: New York, pp. 152–183.
- Kreitler, S./Kreitler, H. (1994a) Who will breastfeed? The cognitive motivation for breastfeeding. International Journal of Prenatal and Perinatal Psychology and Medicine 6: 43– 65.
- Kreitler, S./Kreitler, H. (1994b) Motivational and cognitive determinants of exploration. In: Keller, H./Schneider, K./ Henderson, B. (eds) Curiosity and Exploration. Springer-Verlag: New York, pp. 259–284.
- Kreitler, S./Kreitler, H. (1997) The paranoid person: Cognitive motivations and personality traits. European Journal of Personality 11: 101–132.
- Kreitler, S./Kreitler, H. (In press) Cognitive orientation and health-protective behaviors. Journal of Rehabilitation and Health.
- Kreitler, S./Nussbaum, S. (1998) Cognitive orientation and interest: The motivational understructure for achievement in mathematics. In: Hoffmann, L./Krapp, A./Renninger, K. A./Baumert, J. (eds) Interest and Learning. Institut für die Pädagogik der Naturwissenschaften an der Universität Kiel: Kiel Germany, pp. 377–386.
- Kreitler, S./Shahar, A./Kreitler, H. (1976) Cognitive orientation, type of smoker and behavior therapy of smoking. British Journal of Medical Psychology 49: 167–175.
- Kreitler, S./Schwartz, R./Kreitler, H. (1987) The cognitive orientation of expressive communicability in schizophrenics and normals. Journal of Communication Disorders 20: 73–91.
- LaPiere, R. (1934) Attitudes versus actions. Social Forces 13: 230–237.
- Lévi-Strauss, C. (1979) Myth and Meaning. Schocken Books: New York.
- Lobel, T. (1982) The prediction of behavior from different types of beliefs. Journal of Social Psychology 118: 213–233.
- Lorenz, K. (1965) Über tierisches und menschliches Verhalten. 2 Vol. Piper: München.
- Lorenz, K. (1977) Behind the Mirror. Harcourt, Brace, Jovanovich: New York.
- Lorenz, K. (1981) The Foundations of Ethology. Springer: New York.
- Marom, Z. (1978) Examination of CO Clusters in Children Based on the Theory of Cognitive Orientation. Master’s Thesis, Department of Psychology, Tel-Aviv University, Tel-Aviv, Israel.
- McLean, P. D. (1973) A triune concept of brain and behavior. University of Toronto Press: Toronto.
- Miller, G. A./Galanter, E./Pribram, K. H. (1960) Plans and the Structure of Behavior. Holt, Rinehart & Winston: New York.
- Mithen, S. (1996) The Prehistory of the Mind. Thames & Hudson: London.
- Morris, D. (1969) The Human Zoo. New York: McGraw-Hill.
- Nurymberg, K./Kreitler, S./Weissler, K. (1996) The cognitive orientation of compliance in short and long-term type 2 diabetic patients. Patient Education and Counseling 29: 25–39.
- Plotkin, H. C. (1997) Evolution in Mind. Allen Lane: London.
- Popper, K. (1984) Objektive Erkenntnis. Ein evolutionärer Entwurf. Hoffman and Campe: Hamburg. English orginal: Popper, K. (1972) Objective Knowledge: An Evolutionary Approach. Oxford University Press: Oxford.
- Riedl, R. (1980) Biologie der Erkenntnis. Die stammesgeschichtlichen Grundlagen der Vernunft. 2nd edition. Parey: Berlin, Hamburg. English translation: Riedl, R. (1984) Biology of Cognition: The Phylogenetic Foundations of Reason. John Wiley & Sons: Chichester.
- Riedl, R. (1981) Die Folgen des Ursachendenkens. In: Watzlawick, P. (ed) Die erfundene Wirklichkeit. Piper: München, Zürich, pp. 67–90.
- Riedl, R. (1987) Begriff und Welt. Biologische Grundlagen des Erkennens und Begreifens. Parey: Berlin, Hamburg.
Evolution and Cognition ❘ 96 ❘ 2001, Vol. 7, No. 1
- Riedl, R. (1997) From four forces back to four causes. Evolution and Cognition 3: 148–158.
- Rogers, R. W. (1983) Cognitive and psychological processes in fear appeals and attitude change: A revised theory of protection motivation. In: Cacioppo, J. T./Petty, R. E.(eds) Social Psychophysiology. Guilford Press: New York, pp. 153–176.
- Ronis, D. L. (1992) Conditional health threats, health beliefs, decisions, and behaviors among adults. Health Psychology 11: 127–134.
- Thorpe, W. H. (1974) Animal Nature and Human Nature. Methuen: London.
- Tinbergen, N. (1951) The Study of Instinct. Clarendon Press: Oxford.
- Tipton, R. M./Riebsame, W. E. (1987) Beliefs about smoking and health: their measurement and relationship to smoking behavior. Addictive Behaviors 12: 217–223.
- Westhoff, K./Halbach-Suarez, C. (1989) Cognitive orientation and the prediction of decisions in a medical examination context. European Journal of Personality 3: 31–71.
- Wilson, E. O. (1978) On Human Nature. Harvard University Press: Cambridge MA.
- Wimmer, M. (1995) Evolutionary roots of emotions. Evolution and Cognition 1: 38–50.
- Zakay, D./Bar-El, Z./Kreitler, S. (1984) Cognitive orientation and changing the impulsivity of children. British Journal of Educational Psychology 54: 40–50.
Evolutionary Ethics
Value, Psychology, Strategy and Conventions
1. Introduction
The purpose of this paper1 is to present an example of what moral theory would look like if it took evolutionary theory seriously. In it, I strive to avoid the errors to which so many previous attempts have succumbed. If I succeed in doing so, it is because of my intent merely to show how DAR-WINIAN thinking improves the empirical basis upon which we construct moral theories. At no point do I try to derive foundational moral values either from nature in the broadest sense or from the process of evolution itself. My project is
Abstract
The purpose of this paper is to present an example of what moral theory would look like if it took evolutionary theory seriously. First, I examine briefly the implications that accepting evolutionary theory has for value theory. Secondly, I suggest that an evolutionary perspective can shed new light on moral psychology. Finally, I suggest that an evolutionary perspective clears the way for a moral perspective that focuses on the function and development of HUMEAN social conventions. I strive to avoid the errors to which so many previous attempts at grounding ethics in evolution have succumbed. If I succeed in doing so, it is because my intent is merely to show how DARWINIAN thinking improves the empirical basis upon which we construct moral theories. At no point do I try to derive foundational moral values either from nature in the broadest sense or from the process of evolution itself.
Key words
Ethics, convention, psychology, function, game theory.
based on the widely-held assumption that moral decision making requires both values and facts. Knowing a bit about evolution can help make it clear what the relevant facts are. Here, I do not intend to discuss biological evolution in particular, but evolution in the broadest sense. Evolution takes place wherever there is variation, replication, mutation, and differential survival according to inherited traits. Thus evolutionary theory applies to (at least) biological organisms, certain computer programs, cultures, and ideas.
At the heart of my project lies the claim that moral standards are the products of biological and cultural evolution. Biological evolution has given us brains capable of certain cognitive tasks, and capable of manifesting certain emotions. On top of this biopsychological substrate, cultural evolution has built derstood as regularities of behaviour, sustained by a shared interest in coordination and an expectation that others will cooperate.
Why social conventions? To begin with, as GAUTHIER (1991, p18) argues, we no longer accept the world-view upon which absolutist conceptions of morality depend.2 Thus we no longer have access to plausible notions of inherent and intrinsic value upon which to base ideal moral codes.3 An evolutionary view can only be founded on a subjective and relative theory of value.4 Such a value-theory leads us to ask what moral rules—what social structures—would aid in the pursuit of whatever it is that we value.5 What moral rules have we agreed upon, and what moral rules could we agree upon, in pursuit of what we want? That is, the evolutionary perspective must lead us to focus on the extent to which
Evolution and Cognition ❘ 98 ❘ 2001, Vol. 7, No. 1
a wide range of particular norms and conventions. The field of ethics can be interpreted as being devoted to influencing the course that the evolution of these norms and conventions takes.
The basic claim of this paper is that attention to evolutionary theory should have two main consequences for moral theory. First, attention to evolutionary theory (as it applies to ethics) should draw attention to the field of evolutionary psychology, as a way of enriching our understanding of moral psychology. Secondly, it should draw our attention to the study of social conventions, unmorality is a useful human construct, a necessary artifice.
Our second reason for paying to attention to conventions is that an evolutionary perspective forces us to take an explicitly historical view of our moral standards. Much of moral theory is weakened by the extent to which it fails to appreciate the importance of considering moral agents as historically and socially situated. Moral theory mistakenly assumes, for the most part, that agents (or societies) are in a position of radical choice vis-à-vis their moral principles, or more generally, their social arrangements. Attention to social conventions—actual, existing, working bits of cooperative behaviour—is a way of taking seriously the moral work already done by cultural evolution.
2. Value
We begin by looking at the implications of evolutionary theory for value theory. The most fundamental shift that an evolutionary perspective brings about—or should bring about—in our understanding of value lies in the undermining of what RACHELS (1990) calls the ‘Human Dignity Thesis’. According to the Human Dignity Thesis, humans have special value not shared by nonhuman nature; also according to the Thesis, humans have this special value just by being human, which implies that all human lives have this value to some extent or another. Both of these value claims have wide-reaching implications for ethics. But, according to RACHELS, an evolutionary perspective casts doubt upon the two claims that he calls the ‘twin pillars’ holding up the Human Dignity Thesis. First, the acceptance of evolutionary theory renders unnecessary (as an explanatory hypothesis) the Western religious claim that humans were made in the image of God. Secondly, in situating humans on a developmental spectrum of cognitive capacities, as creatures with capacities different in degree but not in kind from other animals, evolutionary thinking undermines the secular claim that humans are of special value on account of their unique rationality.
Although the theory to which, I argue, the evolutionary perspective drives us is the theory of value assumed by our most well-developed social science—economics—it is also a theory of value that is at odds with most modern moral theories. A full theory of value is beyond the scope of this paper, but the basics—borrowed from David GAUTHIER’s account of value—can be stated briefly. The value theory to which an evolutionary perspective drives us is one that holds that all value is dependent upon the affective relationships of the individual, and is therefore both subjective (as opposed to objective) and relative (rather than absolute).
First, that value is subjective means that the ascription of value is taken to be attitudinal, not observational (GAUTHIER 1986, p25). That is, a thing’s being valuable depends upon some sentient being having a positive attitude toward it. GAUTHIER (1986, p46) puts the point quite simply: “Value… is a measure of preference”. Thomas HOBBES is perhaps the classic exponent of the subjective view of value. HOBBES (1985, p24) writes, “For these words of Good, Evill… are ever used with relation to the person that useth them: There being nothing simply and absolutely so; nor any common Rule of Good and Evill, to be taken from the nature of the objects themselves; but from the Person of the man”. For the value subjectivist then, nothing is ever valuable simpliciter; it is always valuable to someone. According to GAUTHIER (1986, p56), we should decide “whether any adequate account of our experience or environment must refer to objective value” by considering “whether any reference to objective value occurs in the best explanation we can provide for our actions and choices” (emphasis original). The value objectivist, according to (GAUTHIER 1986, p55) “…holds that any adequate account of our experience or our environment must refer to value or to the valuable as being independent of sentient beings and their affections”. GAUTHIER suggests instead that “…the only serious candidate for an explanatory schema for human action is… [one according to which] choice maximizes preference fulfilment given belief. Objective value plays no role in this account” (p56). GAUTHIER continues: “Objective value, like phlogiston, is an unnecessary part of our explanatory apparatus, and as such is to be shaved from the face of the universe by OCKHAM’s razor”.
I would argue that objective value died (or at any rate, should have died) in 1859, with the publication of DARWIN’s The Origin of Species. DARWINIAN theory pulls down the two supporting pillars of the Human Dignity Thesis. Under the scientific world-view with which we are left, objective value is simply an unnecessary explanatory hypothesis.
On the theory of value to which evolutionary thinking drives us, not only is value to be taken to be subjective, but it is also relative. On the relative conception of value, according to GAUTHIER (1986, p50), “…each person has his own good (and bad),
Evolution and Cognition ❘ 99 ❘ 2001, Vol. 7, No. 1
and… the goods of different persons are not parts of a single, overall good”. Note that to hold that value is individualistic in this sense does not imply either ethical or psychological egoism; that is, it does not imply that individuals care only about their own well-being. As GAUTHIER puts it, “A state of affairs that is good for a single individual is then good from the standpoint of each person who prefers or would prefer the enhancement of that individual’s well-being” (p51). (Nor does understanding value as relative necessarily lead to ethical relativism. Moral rules may be shared, even if we differ in our attributions of value.) In opposition to the relative conception of value is the absolutist conception, which holds, according to GAUTHIER that “…values are the same for all persons, or for all sentient beings” (p50). The conception of value as relative fits nicely with the competitive framework of evolutionary theory, according to which individuals may value the welfare of others, but only contingently.
As noted above, most modern moral theories are at odds with this subjective and relative theory of value. Utilitarianism, for example, makes the claim that the happiness of any given individual is a good thing simpliciter; that is, it is to be regarded as a good by every rational being. Thus for Utilitarians, value is subjective but also absolute. KANTIANS, for their part, hold that the respect due to all rational agents finds its source in their inherent value. The KANTIAN theory of value must then be both objective (since it makes no mention of anyone’s preferences or affective states) and absolute (since it holds rational agents to have value for everyone). For a fuller defence of the individualist theory that takes value instead to be both subjective and relative, I refer the reader to GAUTHIER (1986)’s Chapter II section 4.
3. Evolutionary Psychology
How can we tell whether a given rule or standard is consistent with our moral psychology? Empirical investigation—trial and error—is one way.6 But evolutionary considerations can be useful in guiding our empirical investigations. Thus one set of methods that can be used to study the psychological mechanisms behind our moral behaviour is that of evolutionary psychology.7
Evolutionary psychology is predicated on the idea that, contrary to the beliefs of behaviourists such as PAVLOV and SKINNER, the human mind is anything but a blank slate. Rather than seeing it as a generalpurpose computational device, evolutionary psychologists see the mind as a collection of specialpurpose algorithms, each designed (in the metaphorical sense8 ) to solve a particular problem faced during our evolutionary history.
According to evolutionary psychologists, evolution has given us certain cognitive capacities (such as the ability to detect aggression in others, or the ability to remember wrongs) and emotional dispositions (such as the capacity for sympathy and spite) that factor significantly in our moral lives. Investigation of these capacities and dispositions can be informed by evolutionary theory. Knowledge of the sorts of problems that the human brain evolved to solve can lead to hypotheses concerning what sorts of particular cognitive mechanisms and predispositions we can expect to be included in our psychological makeup. Such hypotheses can be tested against anthropological data and against psychological evaluation of test subjects. We can then evaluate particular moral standards in terms of the degree to which they are compatible with known capacities and dispositions. In applying this test, I am proposing the use of an evolutionary perspective to investigate human nature. There is nothing novel about wanting to look at human nature in designing moral systems. The idea here is that our shared nature places limits on what we can learn to do—on how we can learn to behave. The novelty, of course (at least for philosophers) lies in looking to evolutionary theory for a better understanding of that nature.
3.1 Cognitive capacities
What sorts of limits on moral behaviour might be seen in our shared, evolved nature? As one example, COSMIDES and TOOBY (1992) find compelling evidence for a special human aptitude for detecting cheating in situations of social exchange. Similar studies might just as easily find that humans are particularly inept at a particular kind of reasoning. Given such knowledge, it would seem foolish (from an instrumentalist point of view) to propose a moral standard that ignored our aptitudes while relying heavily on forms of reasoning at which we generally do badly.
For example, it might one day be discovered that humans lack the aptitude for categorical reasoning of the sort demanded of us by KANT. That is, it might turn out, upon empirical investigation, that humans are relatively inept at figuring out whether, in a particular case, the maxim by which one acts is one “which [one] can at the same time will that it should become a universal law” (KANT 1959, p39). If such a hypothesis about our cognitive capacities
Evolution and Cognition ❘ 100 ❘ 2001, Vol. 7, No. 1
were validated, I take it that this would be a harsh though perhaps not fatal—criticism of that aspect of KANTIAN ethics (at least from the point of view of anyone who wants to propose useful moral rules).
3.2 Emotional predispositions
Next, let us look at the emotional dispositions with which our evolutionary past has left us. Since ‘ought’ implies ‘can’, it would seem foolish to have a moral rule which demanded behaviours of which we were emotionally (or otherwise) incapable, or nearly so. A case in point: all available evidence points to a general human predisposition to give preferential treatment to friends and family.9 Not only is this tendency apparently universal among human cultures, it also shows up (under the title ‘kin altruism’) in a large number of animal species. Flying in the face of this apparently universal human tendency is the Benthamite claim that “each is to count for one and none for more than one”. This Utilitarian standard requires precisely that my brother not count for more than a stranger in my moral deliberations. I submit that if we want our moral rules to be respected and adhered to, they at least ought not to demand that we forgo strongly ingrained behaviours unnecessarily.
It may be objected that humans display all sorts of nasty predispositions that ought not to be excused simply because they are ‘natural’. I agree, and not simply because I want to avoid committing the naturalistic fallacy. For example, regardless of how ingrained our tendencies toward aggression are, we need to place moral restrictions on aggressive behaviour. Yet in the case both of aggression and of favouritism, the crucial point is to limit socially destructive behaviour. Thus, while our sense of justice demands that we limit favouritism in cases such as the awarding of government contracts, there will be many situations in which preferential treatment of family and friends seems not just forgivable, but appropriate. Likewise, while it is socially imperative to prohibit the killing of innocents, it seems unnecessary to forbid certain other displays of aggression, for example organized sports.
Further, consider the fact that there may be a number of moral rules that could mediate any given social situation. For example, we might determine suitable patterns of economic distribution in one of three ways:
by assigning to each individual the right to some level of quality of life (i.e., a form of Deontology);
by claiming that a certain pattern of distribution maximized the net benefits to society as a whole (i.e., Utilitarianism); or
by claiming that a certain pattern of distribution would be agreed to in a (hypothetical) fair bargaining situation (i.e., Contractarianism).
Each of these rules is in some sense plausible, and each might even point to a similar outcome. So instrumentally, it might not much matter which one we choose. I suggest that if one of them turns out to be psychologically implausible, this can help us choose among these competing moral standards.
4.1 The function of morality
First, we can use game theory to illuminate the function(s) of morality. For example, GAUTHIER (1986) uses game theory to argue that morality is a solution to problems of strategic interaction such as the Prisoner’s Dilemma. Economist Ken BINMORE (1994) argues that morality is a device for choosing among various stable equilibria in multi-person games. (A game is said to be in equilibrium when each player’s choice of strategies is the best response to her opponent’s choice of strategies.)
An example may help to illustrate this point. Imagine a game in which our goal is to agree upon a social contract (in the non-technical sense), which involves negotiating a set of rules for us to live by. We can reduce this problem to its strategic essence by representing it in simplified form as a game. Let us look at the following 2–person game, where x, y, and z are possible actions that each of us could take. If we think of this game as representing negotiation
| You | |||||||
|---|---|---|---|---|---|---|---|
| x | y | z | |||||
| x | 0 | 1 | 3 | ||||
| 0 | 1 | 3 | |||||
| 1 | 2 | 1 | |||||
| Me | y | 1 | 3 | 1 | |||
| z | 2 | 1 | 0 | ||||
| 5 | 1 | 0 |
Figure 1: Choosing a Social Contract. The payoff pairs are {payoff to me, payoff to you}, where payoffs are assumed to correspond to preferences. 5 is best, 0 worst.
toward a social contract, then x, y, and z represent different sets of rules that we could each choose to live by.
In this case, the outcomes {z,x} (at bottom left), {y,y} (centre), and {x,z} (top right) are all ‘equilibria’. That is, for each of those pairs, my choice is my best possible reply to your choice, and your choice is your best possible reply to my choice. For example, if I choose z, you are best off if you choose x, and if you choose x, I am best off if I choose z. Equivalently, we can say that if you and I are already in a situation where I am playing x and you are playing z, neither of us can benefit by unilaterally changing his behaviour. The same is true for the outcome pairs {y,y} and {x,z}.
(This game would be a simple matter of coordination, were it not for the fact that the payoffs in two of the three equilibria are asymmetrical. If the payoffs in all three equilibria were symmetrical, then we would find it easy to agree which equilibrium was best: it would just be a matter of coordinating our efforts.11 As it stands, the example above poses a hard moral problem.)
In the example at hand, we know that we’re better off cooperating than not cooperating, but we disagree over which project to cooperate on (i.e., which social contract to choose). If we are currently both playing y, (i.e., in the centre of the matrix) then I would prefer that we move to the lower–left of the payoff matrix, but I can’t move us there by myself. Changing my behaviour unilaterally would make me worse off. Similarly, you would prefer that we move to the upper–right of the payoff matrix, but you would be worse-off than you are now if you
changed your behaviour unilaterally. The only way either of us can improve our own situation is for both of us to agree on a new equilibrium. BINMORE’s claim is that moral norms are coordination mechanisms that allow us to deal with quandaries such as this. According to BINMORE, moral norms are coordinating conventions for the selection of equilibria. This is the functionalist description of moral standards to which an evolutionary perspective leads us. But how can we endorse a perspective that ‘reduces’ morality to convention? We turn to this question next.
5. Conclusion
I have argued that an evolutionary perspective has three main implications for moral theory. An evolutionary perspective has implications for value theory (suggesting a subjective, relative theory of value), can guide us in interpreting the extent to which various candidate moral rules are consistent with human moral psychology, and can help us understand the function of different moral rules in strategic interaction. And each of these implications pushes us toward a conventionalist understanding of ethics.
A full examination of the significance of social conventions for moral theory is beyond the scope of this paper. I will limit myself to making a few broad, programmatic remarks.
First, we should pay attention to the extent to which existing social conventions structure social interaction and moral choice. Often times, conventions will be so well entrenched (and so psychologically ingrained) that we will not see ourselves as facing decisions at all. Further, existing conventions structure individual moral choices in that it is by reference to existing standards that we justify our actions to others. Justification is not an abstract quality: it is always a matter of justification to someone.15 To the extent to which we are concerned with justifying our actions to members of our own communities, we will need to pay attention to the actual standards—existing, acknowledged social conventions—that apply there, rather than to hypothetical or ideal ones.16 We should never be satisfied to rely upon existing standards; existing social conventions can often be improved upon. But where conventions exist, they will need to be heeded; their contribution must be analysed, and they should be ignored only when a plausible alternative is in sight.
Secondly, we should examine the extent to which conventions should be seen as having normative force. Conventions represent—by definition—working solutions to social problems. Conventions are mutually beneficial, and thus have some motivational force. But conventions will not always be equally beneficial. Some conventions, for example, will be asymmetrical in that they reflect, rather than mitigate, power imbalances between the parties involved. The possibility of asymmetrical conventions is both a challenge to the notion of conventions as normative, and a challenge to the common presumption that the notion of equity is essential to morality.
Finally, we need to investigate the possibility of seeing conventions as a moral technology, and ask whether we can build new conventions to remedy existing social problems. If existing conventions serve a useful function, then perhaps we can take what we have learned about conventions and apply that knowledge in a forward–looking manner. One of the implications of this line of thought is that good conventions make good moral choices easier. It might even be said that the ‘real action’ of morality goes on long before the individual is ever faced with a decision. This points to a need for attention to the building, rather than the following, of moral rules. Is it possible to engineer new conventions? Are there some kinds of conventions which are impossible to
References:
Binmore, K. (1994) Game Theory and the Social Contract, Vol. 1: Playing Fair. Cambridge MA: MIT Press.
- Cosmides, L./Tooby, J. (1992) Cognitive Adaptations for Social Exchange. In: Barkow, J. H./Cosmides, L./Tooby, J. (eds) The Adapted Mind: Evolutionary Psychology and the Generation of Culture. Oxford: Oxford University Press, pp. 163–228.
- Danielson, P. (ed) (1999) Modelling Rationality, Morality,
and Evolution. Vancouver Studies in Cognitive Science, Vol 7. Oxford: Oxford University Press.
- Gauthier, D. (1977) The Social Contract as Ideology, Philosophy and Public Affairs 6: 130–164.
- Gauthier, D. (1986) Morals by Agreement. Oxford: Oxford University Press.
- Gauthier, D. (1991) Why Contractarianism? In: Vallentyne, P. (ed) Contractarianism and Rational Choice: Essays on David Gauthier’s Morals by Agreement. Cambridge: Cambridge University Press, pp. 15–30.
Evolution and Cognition ❘ 104 ❘ 2001, Vol. 7, No. 1
Hobbes, T. (1985) Leviathan. London: Penguin Books. First published 1651.
Kant, I. (1959) Foundations of the Metaphysics of Morals. Translated by Lewis White Beck. New York: Macmillan/Library of the Liberal Arts Press.
MacDonald, C. (1998) Evolutionary Perspectives on Ethics. Encylcopedia of Applied Ethics, Vol. 2. San Diego: Academic Press, pp. 189–196.
MacDonald, C. (2000) Clinical Standards and the Structure of
Professional Obligation. Professional Ethics 8(1): 7–17.
- Norman, W. (1998) Inevitable and Unacceptable?: Methodological Rawlsianism in contemporary Anglo–American political philosophy. Political Studies 46(2): 276–294.
- Rachels, J. (1990) Created From Animals: The Moral Implications of Darwinism. Oxford: Oxford University Press.
- Rawls, J. (1971) A Theory of Justice. Cambridge MA: Harvard University Press.
Zusammenfassungen der Artikel in deutscher Sprache
Jaak Panksepp/Jules B. Panksepp Die sieben Sünden der ‘Evolutionären Psychologie’
In diesem Artikel wird eine fundamentale Kritik an einigen der basalen Voraussetzungen der ‘Evolutionären Psychologie’ (EP) geübt. Die Kritik gründet auf Resultaten ethologischer, ontogenetischer und neurobiologischer Untersuchungen. Der EP wird dabei vorgeworfen, Erkenntnisse dieser Disziplinen nicht zu berücksichtigen und eine ‘Überinterpretation’ der menschlichen Natur vorzunehmen, die völlig von der Idee der ‘inclusive fitness’ beherrscht ist. Vor allem die neurologischen Hintergründe von affektiven und motivationalen Prozessen von Menschen und Tieren stehen dabei im Hintergrund und führen zu einer ungerechtfertigten Ausdehnung des Modulkonzeptes und der damit verbundenen erkenntnistheoretischen Implikationen. Kritisch betrachtet wird auch das Konzept von genetisch determinierten Modulen im Bereich des Neocortex, welche aus der Perspektive der EP ganz bestimmte Funktionen (special purpose functions) erfüllen, deren evolutionäre Hintergründe aus den Überlebensbedingungen des Pleistozäns ersichtlich gemacht werden können.
Dagegen legen die Ergebnisse neurobiologischer Forschungen am menschlichen Gehirn bzw. am Säugergehirn ganz allgemein nur die Existenz von subkortikalen Modulen nahe. Im Bereich ‘höherer’ Hirnregionen erscheint aus neurobiologischer Perspektive nur die Annahme der Existenz eines Sprachmodules als angebracht.
Der Neocortex wir im Gegensatz zur EP eher als ‘Generalist’ interpretiert, d.h. ein auf keine spezifischen Funktionen hin evoluiertes Organ, welches ontogenetischen Einflüssen und Anforderungen gegenüber in hohem Ausmaß offen ist.
Die sieben Sünden der EP werden in folgenden Bereichen geortet:
- Überbetonung der pleistozänen Umgebungsbedingungen, welche sich formend auf menschliches Sozialverhalten ausgewirkt haben sollen
- Extreme Zentrierung der EP auf die menschliche Spezies
- Adaptationismus
- Massive Modularität
- Vermischung von Emotion und Kognition
- Das Fehlen glaubwürdiger neuronaler Perspektiven
- Anti-organische Tendenzen bzw. der Mythos vom computationalen Repräsentationalismus
Ullica Segerstråle
Fortgesetzte Konflikte um Modelle und die Realität
Die Schlacht um die ‘gute Wissenschaft’ die im Rahmen der Kontroverse um die Soziobiologie geschlagen wird setzt sich nun im Bereich der Neurowissenschaften fort. Wissenschaftliche ‘Realisten’ prallen dabei mit eher theoriefreudigen ‘Modellierern’ zusammen. Aus der Sicht von PANKSEPP/PANKSEPP sollten die Vertreter der EP – um entsprechend wissenschaftlich und empirisch fundiert zu sein – die Anatomie und die evolutionären Hintergründe des menschlichen Geistes berücksichtigen.
Außerdem ähnelt die PANKSEPP’sche Kritik an den kognitiven Neurowissenschaften den ethologischen Einwänden an der Soziobiologie, die nicht nur die ultimaten Funktionen von Verhalten in Betracht ziehen, sondern ebenso die Entwicklung, die Evolutionsgeschichte und proximaten Ursachen des Verhaltens. Dies mag auf einen potentiellen Konfliktbereich zwischen Ethologie und EP hinweisen.
Dabei sind PANKSEPP/PANKSEPP in guter Gesellschaft anderer kritischer Positionen, wie den an Entwicklungsprozessen orientierten Wissenschaftlern, die davon ausgehen, daß die Modularität im erwachsenen Gehirn nicht auf eine vorgegebene Bereichsspezifität hinweist, sondern aus epigenetischen Prozessen resultiert.
In der Tat könnte gegenwärtig eine neue interdisziplinäre Allianz entstehen, indem die Kritiker von EP und Soziobiologie auf ihrer Suche nach einem eher ganzheitlichen und pluralistischen Paradigma die Ethologie entdeckt haben.
Evolution and Cognition ❘ 106 ❘ 2001, Vol. 7, No. 1
Schließlich sollte man nicht vergessen, daß die EP ein sich entwickelndes Forschungsprogramm darstellt und eventuell eine Annäherung von kognitiver und affektiver Neurowissenschaft zustande kommt. Ein möglicher Vermittlungsbereich könnte das Feld der nonverbalen Kommunikation sein.
Linda Mealey ‘La plus ça change…’: Antwort auf eine Kritik der EP
PANKSEPP/PANKSEPP stehen nicht alleine in ihrer Kritik an der ‘Evolutionären Psychologie’. Die meisten Kritiker, wie auch PANKSEPP/PANKSEPP sind dabei nicht gegen Evolution oder Psychologie bzw. gegen die Verwendung evolutionärer Konzepte in der Psychologie eingestellt. Die Kritik richtet sich dabei vor allem gegen eine bestimmte, bedeutsame Gruppe von der EP nahestehenden Wissenschaftlern, die eine gemeinsame Haltung zu zentralen Kernproblemen einnehmen.
Ich teile die Bedenken von PANKSEPP/PANKSEPP hinsichtlich der Rigidität und doktrinären Einschränkungen die in manchen Versionen der EP gegeben sind. Jedoch ist diese Debatte nicht neu und ich glaube, daß sie, wie auch andere wissenschaftliche Kontroversen durchaus förderliche Eigenschaften haben.
Carlos Stegmann Kommentar zu Panksepp/Panksepp
PANKSEPP/PANKSEPP behaupten, daß die EP – um innerhalb des Rahmens wissenschaftlicher Forschung zu verbleiben – mit den Erkenntnissen der Neurowissenschaften und der Hirnforschung übereinstimmen muß. Dieses Argument wird im Kommentar erweitert. Die menschliche Spezies stellt eine integrale Einheit dar, die aus unterschiedlichen Schichten besteht. Dazu gehören sowohl die Schichte des soziokulturellen Verhaltens von Gruppen, wie die Schichte des individuellen Verhaltens und die Schichte der neuronalen Mechanismen. Jeder Versuch einer Erklärung der menschlichen Natur, inklusive der Psychologie muß daher interdisziplinär ausgerichtet sein.
Psychische Phänomene müssen aus der soziokulturellen Schichte heraus erklärbar sein, wie beispielsweise diejenigen die Friedrich HAYEK in Bezug auf die ‘extended order’ der menschlichen Gesellschaft entdeckt hat. Ebenso müssen diese Gesetze
auf die neurophysiologischen Gesetze rückführbar sein, die auf dieser Schichte anzutreffen sind.
Nur derartige interdisziplinäre Ansätze stellen eine Kompaß bereit anhand dessen man durch das dunkle Meer des menschlichen Gehirns und der zugehörigen Geistphänomenen navigieren kann.
Russell Gardner, Jr. Affektive Neurowissenschaft, Psychiatrie und Soziophysiologie
PANKSEPP/PANKSEPP betonen in anerkennenswerter Weise daß die EP nicht die stammesgeschichtlich alten Bereiche des menschlichen Gehirns in Betracht zieht. Die EP vermischt subkortikale emotionale Faktoren mit Kognitionen, die im Verlauf der Evolution des Menschen verstärkt wurden. Weiters beachtet die EP die kommunikativen und sozialen Bereiche der menschlichen Entwicklung zu wenig, obwohl neue Arbeiten deutlich machen, daß ein signifikanter Anteil der Varianz der Hirngröße von Primaten mit der jeweiligen Gruppengröße zusammen hängt. Jaak PANKSEPPs frühere Arbeiten über emotionale Kommunikation könnten dabei zur Entwicklng einer grundlegenden psychiatrischen Wissenschaft beitragen, die derzeit leider kaum existiert.
In Übereinstimmung mit PANKSEPP steht die Forderung nach einem basalen, wissenschaftlichen Rahmen für die Psychiatrie mit dem Titel Soziophysiologie, welche sowohl die Ähnlichkeiten als auch die Gemeinsamkeiten des Menschen mit anderen Tieren untersucht.
Gerhard Meisenberg Grade der Modularität
Die Unterscheidung zwischen kognitiven und modularen Funktionen, wie sie ursprünglich von FO-DOR (1983) vorgeschlagen wurde ist gültig. Jedoch werden zwei Formen modularer Funktionen unterschieden.:
‘Primäre’, genetisch determinierte Module, die überwiegend subkortikal vorliegen und hauptsächlich Motivation (z.B. Konditionierung von Angst) bedingen.
‘Modularisierte’ Funktionen, die vor allem kortikal vorliegen und durch wiederholten sensorischen und /oder kognitiven Input geformt werden. Diese Funktionen sind vor allem für sensorische Analyse und motorische Kontrolle verantwortlich (z.B. Sprache).
Evolution and Cognition ❘ 107 ❘ 2001, Vol. 7, No. 1
Die strikte Unterscheidung zwischen genetisch determinierten subkortikalen Schaltkreisen und nicht modular organisierten kognitiven Funktionen, wie sie von PANKSEPP/PANKSEPP vorgeschlagen wird, wird zurückgewiesen. Ebenso wird das von der EP vertreten Modell des ‘Schweizer Armeemessers’ als zu vereinfachend abgelehnt.
Im Gegensatz dazu wird behauptet, daß viele Hirnfunktionen (inklusive der kognitiven Funktionen) ‘Mehr-Zweck’–Mechanismen darstellen. Diese ‘guns for hire’ stehen in Form von spezialisierten Modulen zur Verfügung um adaptives Verhalten zu erzeugen.
Es wird behauptet, daß das nützlichste Konzept ein ausgedehntes ethologisches Modell darstellt, in dem spezialisierte Gehirnsysteme entweder sensorischen oder kognitiven Input verwenden und damit entweder Verhalten oder kognitiven Output erzeugen.
Ian Pitchford Keine Evolution. Keine Kognition
Obwohl sowohl Natur- wie auch Geisteswissenschaftler jeglichen Glauben an unterschiedliche materielle und immaterielle Substanzen leugnen würden, werden gegenwärtige Debatten größtenteils mit einer Terminologie geführt, die antiken griechische Philosophen wohl sehr vertrau gewesen wäre. Dabei werden menschliche Merkmale in göttlich – transzendente und tierisch – körperliche unterteilt. Die modernen Fassungen dieser Unterteilungen lauten dann rational, kognitiv, diskursiv, autonom und kreativ – für den göttlichen Anteil, sowie emotional, instinktiv, determiniert etc. für den tierisch – körperlichen Teil.
Dieser grundlegende Dualismus erhält die, in fast allen Diskussionen über den Menschen anzutreffenden drei zentralen Dichotomien Körper/Geist, Kognition/Emotion und Natur/Kultur aufrecht. Das Bemühen der neuroevolutionären Psychobiologie das Gehirn in einen determinierten affektiven Teil und einen plastischen, nicht direkt funktionsgebundenen Neocortex zu unterteilen, indem Konzepte wie ‘exaptation’, Emergenz und dgl. mehr verwendet werden steht genau in dieser quasitheologischen westlichen philosophischen Tradition. Dieser dualistische Ansatz stellt keine schlüssige Basis dar, von der ausgehend die EP kritisiert werden kann.
Scott Atran Der ‘Fall’ Modularität: Sünde oder Erlösung?
Die These von evoluierten kognitiven Modulen beruht auf unterschiedlichen, konvergierenden Beweisen: functional design (Analogie), Ethologie (Homologie), Universalität, Unabhängigkeit von Perzeptionen, spezifische Pathologie (zerebrale Störungen), Hemmungswiderstand (Hyperaktivität), Leichtigkeit der kulturellen Übertragung.
Zwar mag keiner der Einzelfaktoren notwendig sein, jedoch ist die Datenlage zwingend und eindeutig für Bereiche wie beispielsweise ‘folk biology’ und ‘folk psychology’.
Im Gegensatz dazu wird die zentrale These von PANKSEPP/PANKSEPP durch keinerlei empirische Hinweise gestützt. Diese These geht davon aus, daß modulare Strukturen – wie sie von der EP (neben zahlreichen kognitiven Psychologen und Entwicklungspsychologen) interpretiert werden – epigenetische Produkte subkortikaler Emotionen (die bei allen Säugetieren vorkommen) und neokorticaler unspezifischer ‘Mehr-Zweck’–Operationen sind.
Keine einzige bedeutsame empirische Entdekkung im Bereich Sprache oder anderer höherer kognitiver Funktionen ist bei den Untersuchungen zu den evolutionären Ursprüngen und Funktionen offen geblieben.
Jaak Panksepp/Jules B. Panksepp: Weiter Kritik an der EP: 7 Sünden für 7 Sünder – plus/minus 2
Die vielfältigen, von den Kommentatoren behandelten Problembereiche ließen zwei zentrale Fragen offen: eine Debatte über Modularität sowie die Evolution von Sprache. Dahingehend bietet eine neokortikale – entwicklungsbezogene – systemische Perspektive, zusammen mit einem Verständnis der basalen emotional – motivationalen Systeme (die wir mit anderen Säugetieren teilen) mehr Einsichten in die Formen intrinsischen Funktionierens menschlicher Gehirn/Geist Prozesse als jegliche Variante kortikaler Modularität.
Es erscheint sehr unwahrscheinlich, daß die Entwicklung höherer kortikaler Funktionsbereiche, welche Sprache bedingen, ohne zugrundeliegende, subkortikale Systeme abgelaufen ist. Obwohl Menschen ‘kommunikative Instinkte’ bzw. über ’Anla-
Evolution and Cognition ❘ 108 ❘ 2001, Vol. 7, No. 1
gen zum Spracherwerb’ verfügen können ist die Existenz von sog. ‘Sprachmodulen’ nicht belegt. Dahingehend erscheint es angebracht, daß die Vertreter der EP neurowissenschaftliche Erkenntnisse in ihre Überlegungen einbeziehen.
SEGERSTRÅLE – eine historische Perspektive: SEGERSTRÅLE stellt diese Kontroverse in den aktuellen historischen Kontext. Wir stimmen dabei mit ihr überein, daß die Ethologie in die Fundamente psychologischen Denkens einbezogen werden soll. Wir stimmen nicht mit ihrer Behauptung überein, daß ‘wirkliche Wissenschaft’ nur auf einer ‘anatomisch–physischen Ebene’ gemacht wird, sondern räumen theoretischen Perspektiven ebenso einen zentralen Stellenwert ein.
SEGERSTRÅLE bemerkt zutreffend, daß unsere Bemühungen gegen einen wissenschaftlichen ‘Tribalismus’ gerichtet sind, der eine Fülle von Datenmaterial über ‘nonverbale Kommunikation’ sowie die zugrundeliegende neurophysiologische Basis aus dem tierischen Verhaltensbereich völlig mißachtet. Wir wenden uns gegen die anthropozentrische Perspektive der EP und gehend dabei davon aus, daß zahlreiche, von der EP postulierte, in höheren kortikalen Bereichen angeordnete Module bereits in subkortikalen Arealen (die sich ebenso in anderen Tiergruppen finden) vorliegen. Dabei ziehen wir nicht in Zweifel, daß es innerhalb des Kortex beträchtliche sensorische und motorische Spezialisierungen gibt (inklusive diverser Gesichts-, Geruchs-, Berührungs- und Stimmlagen–Erkennungsdispositionen). Jedoch sind diese Dispositionen weder soziobiologisch (im eigentlichen Wortsinn) noch nur beim Menschen anzutreffen.
Wir stimmen SEGERSTRÅLE dahingehend zu, daß es an der Zeit ist “to relax the modularity idea”.
MEALEY – Was ist in einem Wort? MEALEY gibt eine ausgewogene Übersicht über die grundlegenden und einige spezielle Argumente. Dabei stimmen wir nicht mit ihrer Verteidigung der modularen Sichtweise überein.
Neben allen anderen ‘Sünden’ ist dabei die ‘Spezieszentrierte Perspektive’ zentral. Wir vermissen dabei die Integration neurowissenschaftlichen Datenmaterials in die akademische Psychologie – ohne diese Integration kann die Psychologie niemals eine ‘grundlegende Wissenschaft’ werden.
Gegen den Modularitätsbegriff wie ihn MEALEY verwendet wenden wir ein: er vermittelt ein unzureichendes und vereinfachendes Bild der Hirnfunktionen, die als zusammengeschlossene (encapsulated) Informationsverarbeitungssysteme interpretiert werden. Dabei werden die zahlreichen Wechselwirkungen innerhalb des Gehirns völlig mißachtet. Einen ähnlichen Fehler machte auch die frühe Neurowissenschaft mit dem Postulat von sog. ‘Zentren’ – welches bald nicht mehr verwendet wurde.
Der Modularitätsbegriff stellt im Bereich der Psychologie kein neues und sinnvolles Konzept dar. Dagegen scheint es sinnvoll etwa den Begriff des ‘Triebes’ wieder einzubringen, da dieser eher den Erkenntnissen neurowissenschaftlicher Untersuchungen entspricht.
STEGMANN – eine philosophische Perspektive: Aus einer philosophischen Perspektive heraus stimmen wir mit STEGMANN überein, jedoch aus der wissenschaftlichen Perspektive heraus üben wir Kritik daran. Derartige holistische Perspektiven sind oft einer systematisch – experimentellen Analyse entgegengesetzt.
Ebenso wenden wir uns gegen die Annahme, daß komplexere Schichten auf darunterliegende Systemebene reduzierbar sein sollen. Dieser Form des Reduktionismus stellen wir den modernen philosophischen Begriff der ‘supervenience’ entgegen. Wir wären sehr daran interessiert zu erfahren, inwieweit STEGMANNs Ideen empirischer Überprüfung zugeführt werden könnten.
GARDNER – eine neuropsychiatrische Perspektive: GARD-NER geht von der – im Bereich der Psychiatrie besonders dringlichen Notwendigkeit der Integration unterschiedlicher Perspektiven hinsichtlich Gehirn/Geist Phänomenen aus. Nach wie vor werden diese Phänomene getrennt behandelt und die Notwendigkeit diverser Brückenschläge ist eine der großen gegenwärtigen Herausforderungen.
Es ist eines der wesentlichen Anliegen des Seniorautors die tiefen biologischen Muster psychiatrisch/ emotionaler Störungen aufzuzeigen. Überaus hilfreich ist dabei die Einbeziehung von Datenmaterialien aus tierischen Verhaltensanalysen. Die dabei vorgefundenen basalen emotionalen Systeme, die wir mit allen Säugetieren gemeinsam haben erweisen sich dabei besonders für die Psychiatrie von großem Wert. Die Entstehung der neuro-psychoanalytischen Bewegung erscheint als ein hoffnungsvoller Schritt in diese Richtung.
MEISENBERG – Modularität ist am Leben und guter Dinge – angeblich. Meisenberg scheint mit den größten Teil unserer Überlegungen übereinzustimmen
Evolution and Cognition ❘ 109 ❘ 2001, Vol. 7, No. 1
und unternimmt eine weiterreichende Analyse kortikaler Funktionen.
Wir selbst zweifeln nicht daran, daß große Teile des Neokortex – als Folge von Erfahrungen spezialisiert sind. Jedoch stellen wir die Existenz soziobiologisch begründeter, kognitiver Module in Zweifel. Es gibt keinerlei Hinweise darauf, daß sich modularisierte, kognitive Adaptationen im Neokortex – im Verlauf der letzten 2 Jahrmillionen entwickelt haben. Wir gehen davon aus, daß dasjenige, was sich in diesem Zeitrahmen entwickelt hat eher unspezifische, d.h. nicht spezialisierte Funktionsbereiche waren, die erst unter Umwelteinflüssen und mit subkortikaler Beteiligung modularisiert wurden.
Wir stimmen mit MEISENBERGs Überlegungen hinsichtlich der Entwicklung des Lächelns überein. Jedoch stimmen wir ihm nicht zu, daß die relevanten neurologischen Strukturen vor allem im cingulären Kortex angeordnet sind. Demgegenüber nehmen wir an, daß auch noch tieferliegende Gehirnregionen am Lächeln beteiligt sind.
In striktem Widerspruch stehen wir zu MEISEN-BERGs Behauptung daß das Datenmaterial welches sich auf die subkortikale Ebene bezieht nicht so solide und fundiert ist wie wir das behaupten.
PITCHFORD – die Herausforderung eines Insiders. PITCH-FORD bezieht sich vor allem auf eine der von uns postulierten Sünden der EP – die Suche nach adaptiven Modulen Bereichen, in denen jedoch nur nicht funktionsgebundene Operationen vollzogen werden. PITCHFORD thematisiert dabei vor allem die Sprachfunktion und die Rolle von Entwicklungsprozessen.
PITCHFORDs Behauptung die Autoren hätten die zentralen Argumente der ‘developmental systems theory’ nicht entsprechend berücksichtigt wird zurückgewiesen, da vor allem der Seniorautor dazu zahlreiche Publikationen vorgelegt hat. Dabei stimmen wir größtenteils mit den Argumenten der ‘developmental systems theory’ überein – ein Kritikpunkt jedoch richtet sich gegen jene überzeichnete Argumentation in der von ‘Ontogenie von Information’ die Rede ist.
Leider geht PITCHFORD nicht auf unsere Kernargumente ein.
Noch einmal weisen wir darauf hin, daß der gefährlichste Ort der Suche nach Modularität derjenige von hoch spezialisierten neokortikalen Arealen im Gehirn erwachsener Menschen ist. Wir sind der Meinung, daß sich die allgemeinsten Funktionsprinzipien eher auf subkortikalem Niveau finden.
Atran – noch einmal das ‘Sprachmodul’. ATRAN liefert eine heftige Verteidigung der Modularitätsidee und bezieht sich dabei vor allem auf die Sprache. Das Zentralproblem dabei kann folgendermaßen formuliert werden: beinhalten die Gene die Rudimente unserer einzigartigen menschlichen Sprache oder helfen diese Gene mit komplexe Laute zu erzeugen, die mit dem enormen Speicherraum des Kortex interagieren und als Endprodukt Sprache generieren.
Nach unserem Dafürhalten ist es nicht notwendig die Existenz von genetisch fixierten Sprachmodulen anzunehmen. Sprache entwickelt sich aus dem Zusammenwirken von subkortikalen Hirnstrukturen mit dem Neokortex.
Wir stimmen auch nicht mit ATRANs ‘kortikal – kognitivistischer Sichtweise’ überein, nach der keinerlei Befunde aus dem subkortikalen Bereich Einsichten in das Funktionieren kognitiver Strukturen vermitteln konnte. Die Bedeutung und der Einfluß subkortikaler Aktivitäten wurde und wird nach wie vor weitgehend unterschätzt.
Shulamith Kreitler Eine evolutionäre Perspektive auf kognitive Orientierung
Der Beitrag beschäftigt sich mit den evolutionären Aspekten der kognitiven Steuerung des Verhaltens. Aufgrund empirischer Befunde und der eindrucksvollen Entwicklung von Kognition im Verlauf der Evolution kann davon ausgegangen werden, daß die kognitive Verhaltenssteuerung in Bezug auf Überleben eine Rolle spielt. Eine Theorie kognitiver Orientierung – auf dem Niveau des Menschlichen – wird als grundlegender Rahmen für eine evolutionäre Analyse präsentiert. Die großen Vorteile einer solchen Theorie bestehen in einer umfassenden Beschreibung der zwischen Input und Output wirkenden Prozesse, für die es vielfache empirische Belege gibt. Die wichtigsten evolutionären Aspekte, die besprochen werden, betreffen die Unterscheidung zwischen Motivation und Kognition, die Entwicklung von Bedeutung sowie die Entwicklung von Handeln, in jedem Fall unter besonderer Berücksichtigung der Rolle von Kognition.
Evolution and Cognition ❘ 110 ❘ 2001, Vol. 7, No. 1
Mac Donald Evolutionäre Ethik: Werte, Psychologie, Strategie und Konventionen
Absicht dieses Artikels ist es deutlich zu machen, welche Auswirkungen eine strikte Anwendung der Evolutionstheorie auf eine Theorie der Moral hat. In einem ersten Schritt werden die Implikationen der Evolutionstheorie für die Werttheorie und das moralische Verhalten aufgezeigt. Danach wird der Versuch unternommen eine evolutionäre Perspektive hinsichtlich des Problems der Entwicklung sozialer Konventionen darzustellen. Moralische Standards werden dabei als Produkte der kulturellen und der biologischen Evolution begriffen, wobei basale moralische Werte weder von der Natur noch vom Evolutionsprozeß selbst abgeleitet werden. Die Anwendung des DARWINISTISCHEN Denkens erfolgt dabei vor allem auf die empirische Basis, auf der aufbauend moralische Theorien entwickelt werden.
Bezug genommen wird dabei auch auf die ‘Evolutionäre Psychologie’ welche viel zum Verständnis moralischen Verhaltens beitragen kann.
Weiters erweist sich die Untersuchung sozialer Konventionen als zentrales Element. Konventionen werden dabei als Regularitäten des Verhaltens interpretiert. Diese werden durch mit anderen Menschen geteilte Interessen aufrechterhalten wobei die Erwartung der Kooperationsbereitschaft zugrunde liegt.
Comments on Jaak Panksepp and Jules B. Panksepp’s Paper “The Seven Sins of Evolutionary Psychology”
HAVE READ WITH GREAT interest Jaak and Jules B. PANKSEPP’s paper “The Seven Sins of Evolutionary Psychology”, whose main thesis meets with my full agreement, to wit, that evolutionary psychology should fully consider, and be consistent with, the findings of cognitive neuroscience and brain research, to “remain on the shores of sound scientific inquiry”. I
I would like to extend this thesis: any discipline must fully consider, and be consistent with, other, related disciplines, to produce credible results. This applies especially in the realm of human nature and behavior, which are the subject of many disciplines, among them sociology, political and legal science, behavioral biology and comparative behavioral research, moral
science, and epistemology, in addition to the two disciplines that are the subject of the paper.
In a paper which I have submitted to Evolution & Cognition, “The Human Behavior Instinct. How Decisions for Action are Reached. An Interdisciplinary Theory of Human Behavior”, I have posited the following general postulate to account for the required interdisciplinarity:
Abstract
The commented paper states that, “to remain on the shores of sound scientific inquiry” evolutionary psychology should be consistent with the findings of cognitive neuroscience and brain research. This argument is extended in the present comments. The human species is an integrated natural entity composed of strata, including the strata of sociocultural group behavior, of individual behavior, and of neural mechanisms. Events or phenomena affecting the entity are one–and–the–same in all and each of the strata, and thus any factually correct explanation of human nature, including psychology, must be interdisciplinary in scope. Psychological phenomena must be explainable in terms of reductions of laws which govern the sociocultural stratum, such as those discovered by Friedrich HAYEK related to the extended order of human society, and be reducible in turn to the mechanisms that operate in the neurophysiological stratum. The comments conclude that only such an interdisciplinary approach can provide a compass to navigate in the dark sea of human brain and mind phenomena.
Key words
Interdisciplinarity, psychology, cognition, human behavior, evolution, neuropsychology.
Evolution and Cognition ❘ 20 ❘ 2001, Vol. 7, No. 1
“Living beings, including Homo sapiens, are integrated natural units or entities (systemic wholes) composed of strata, from the atomic-particle to the socio–cultural strata. Stratum-specific natural laws apply in each stratum, but events or phenomena, which affect the entity, are one–and–the– same event or phenomenon in all and each of the strata. Since the event or phenomenon is one– and–the–same, there cannot be any ‘causation’ between strata, but the natural laws which apply in each stratum must be reducible to the laws which apply in the hierarchically lower strata; the idea of natural law only makes sense in this context. Another consequence is that all the strata of an integrated unit evolve together; there can be no operate efficiently in groups. Such effects may emerge more rapidly via group selection than by individual selection.” (My Italics.)
But the groups and the individuals that compose them, and the behavioral phenomena, which affect both groups and individuals, are one–and–the–same thing. However, the natural laws, which govern the behavior of groups, must be reducible to the natural laws, which govern the behavior of the individuals that compose them. This applies to all the integrated natural entities of living beings, but is especially relevant for the natural entity homo sapiens. For this entity possesses an upper stratum of socio–cultural phenomena, which the other entities do not, excepting may be some rudiments in the societies of great apes. And here another critique may be added to the objection that evolutionary psychologists (and sociobiologists as well) do not sufficiently consider the relevance of brain research in their inquiries. The phenomena investigated by psychology occur in the individual behavioral stratum of the human entity, which is interposed between the stratum of neurophysiological phenomena, and the stratum of group behavior in human society. This stratum has been the object of Friedrich A. HAYEK’s work (1982, 1988). HAYEK created the concept of the spontaneous evolutionary formation of the general laws of conduct that govern the extended order of human society. Psychology, especially evolutionary psychology, should consider the relevance of these findings at the level of human society, and be able to provide their reduction to individual human behavior. Besides, individual behavior is also the object of comparative behavioral research, (LORENZ 1973, 1978; RIEDL 1988), and of moral science (Immanuel KANT 1904)1 . It would also be worthwhile to consider, for finding out how the brain/mind works, the conclusions of epistemology, especially those related to analytic and synthetic cognition (CARNAP 1995). Psychologists, however, seem to be working almost exclusively within the limits of their own scientific realm.
In the final part of the commented paper (p125) the following question is posed:
“How do we incorporate the strikingly relevant evidence culled from our fellow animals into the exceedingly anthropocentric modes of thought that characterize so much of present day psychology?”
I do not think that psychology’s modes of thought are “exceedingly anthropocentric”. Psychology is, after all, a human science; its object is the explanation of human behavior. Psychology, to be effective, must clearly incorporate in its research “the underlying brain processes shared by all mammals”
(p125). But it must also incorporate the findings of comparative behavioral research, of moral science, and of epistemology, and be effective in providing explanations (reductions) of the socio–cultural behavior of groups. The referred question is followed immediately by another one:
“How shall we accept our animal heritage without demeaning our vast intelligence?”
But our “vast intelligence” is included in our “animal heritage”! As conclusively demonstrated by Adolf HESCHL in his book “The Intelligent Genome” (1998), the only possible mechanism of creating information is the random mutation–and–selection mechanism of evolution, the genome being its sole repository. All existing information is innate, and this information includes the structural and operational ‘design’ of those vast brain and nerve systems of our species, where our “vast intelligence” originates.
The general overall proposition of the commented paper is that explanations of human behavior should arise from the interaction between the objective cognitive abilities which reside in the massive general-purpose human neocortex, and the emotional and motivational systems which reside in the very old subcortical brain areas, and not, as proposed by evolutionary psychology, from any “finegrained molding of special-purpose socio-affective mechanisms”, or “new and refined emotional modular functions that would have emerged in the human brain/mind during the past several million years of human brain evolution”. I quote several passages of the paper, related to this proposition, to which many more may be added:
On p108: “…we must remember to be especially cautious in ascribing discrete special-purpose functions to brain association areas that appear at birth to be largely general-purpose ‘computational’ devices.
On p110: “The organization of the neocortex, although still constrained by many unknown genetic rules, … may be much more of a general-purpose computational device than modern evolutionary psychologists have been willing to concede”, and “The possibility is remote … that many unique and detailed epistemological engravings of sociobiological strategies (i.e., modules) exist within the human neocortex.”
On p113: “It seems to us that much of brain evolution during the Pliocene and Pleistocene eras was based upon the rapid expansion of general-purpose cortico-computational space … rather than on any fine-grained molding of special-purpose socio-affec-
Evolution and Cognition ❘ 21 ❘ 2001, Vol. 7, No. 1
tive mechanisms. Most special-purpose mechanisms in the brain, of which there are many in subcortical regions, evolved long before humans emerged as contenders for the top ‘predator’ position in the feeding hierarchy.”
On p122: “The existence of various emotional and motivational feelings, along with general-purpose learning systems, can provide practically all the types of adaptive behavioral strategies that have been discussed by evolutionary psychologists.”
And on p125: “In our estimation, the type of psychological functions that evolutionary psychologists speak of, arise largely from the utilization of very old emotional capacities working in concert with newly evolved inductive abilities supported by the vast general-purpose neocortical association areas.”
I fully agree with all those statements. But if the ‘modular’ explanation of evolutionary psychology is not accepted, what other explanation can be proposed? What the paper proposes is a general approach, but it does not suggest a possible concrete explanation that would substitute for the ‘modules’ of evolutionary psychology. As I said at the beginning, a more thorough explanation can only arise from a comprehensive interdisciplinary approach, in which all the strata of the integrated natural entity Homo sapiens are considered. The main concepts, which govern such an interdisciplinary approach, may be put as follows:
The phylogenetic evolution which culminated with the appearance of homo sapiens must have included several instances of the phenomenon ‘fulguration’ (LORENZ 1973, 1978), whereby, through the combination of various elements, a new category of existence comes into being, the properties of which could not have been predicted from the properties of the composing elements. The new entity now possesses a new upper stratum, but it is still the integrated whole of all the strata that compose it, from the atomic-molecular to the spiritual. Certain natural laws act in every stratum, and explanations or reductions must be considered possible. The new species is distinguished, chiefly, by the possession of a new cognitive apparatus. This apparatus operates with two different mechanisms or processes:
The mechanism or process of objective cognition, usually called reason, which contains the analytic faculties which permit to derive synthetic knowledge about reality (CARNAP 1995), combined with the faculty of language and a greatly expanded memory, and which superseded (but still includes), as a result of the fulguration, the ratiomorph faculties of the pre-human ancestor;
The mechanism or process of moral cognition, usually called conscience, which relates objectively known options for action to, also objectively known, values, which in turn are related to certain feelings which the apparatus generates to make us act (DAMASIO 1994), and which superseded (but still includes), as a result of the fulguration, the action-governing instincts of the pre-human ancestor.
This agrees with the following statement of the commented paper (p116):
“…the classic distinction between emotional and cognitive processes is sustained by abundant data indicating that the two can be dissociated functionally and anatomically…”.
At the behavioral psychological level, human decision making is based on beliefs. In objective cognition, a belief is the conviction of the truth of a statement, that is, of the correspondence of the statement with the objective fact that it intends to describe. The objective fact may refer either to the properties of an object, as in scientific discovery, but also, e.g., in engineering design, in business administration, and in many other practical fields, to the objective means and ways to attain a purpose, in a range that extends from the many everyday decisions for action we take on the spur of a moment, to the elaborate plans based on PERT or CPM methods in large enterprises. In moral cognition, a belief is the conviction of the rightness of instrumental and end values, i.e., moral general norms of conduct, and institutions, which apply generally to all, like democracy or socialism. These moral convictions constitute a general framework for our decisions for action.
A belief is actually the feeling of the emotions which govern our decisions for action, and which, as said in the commented paper, “arise largely from the utilization of very old emotional capacities” in the ancient subcortical structures of the brain, and is attached, or affixed, to objectively known concepts in “the vast general-purpose neocortical association areas”.
In objective cognition a scientist is convinced of the truth of his or her theory, and an executive of the correctness of his or her approach to a practical problem. However, although both hold on firmly to their beliefs, they will change them if well-argued fundamental criticism makes this necessary. Thus, evolutionary psychologists may change the approach to their subject as a result of the PANKSEPP’s critical observations, or an engineer his manual methods for
Evolution and Cognition ❘ 22 ❘ 2001, Vol. 7, No. 1
computer-aided calculations. Objective beliefs are thus always reversible, as objective cognition evolves by prediction and critique. It can be argued that inclusive-fitness evolution has developed the reversible feature of objective beliefs to govern collaboration between individuals and groups, in adaptation to the ‘prediction–and–critique’ feature of this cognitive field.
In moral cognition, however, people hold on adamantly to their convictions in the rightness of instrumental and end values. They will not change their ways of behavior and institutions, no matter what kinds of arguments are employed to this end. This is an historical fact. Early Christians preferred martyrdom, Giordano BRUNO the stake, SOCRATES the hemlock, so many patriots torture and death, to renouncing their beliefs. Moral values are not objectively demonstrable. To try to do so is the naturalistic fallacy. This fallacy can be expressed in objective terms through the assessment that any change of the instrumental and end values (general laws of behavior and institutional aims which govern the extended order of society) will provoke so many intricately interwoven direct, indirect, and retroactive effects, that their analysis would involve combinatorial numbers of astronomical dimensions, and thus make a prediction of their overall effects on the structure of the extended order all but impossible. It is only ex–post–facto, that is, after the practical application of the new value for a period of time, that its appropriateness or inappropriateness can be dem-
onstrated. Moral beliefs are thus irreversible, as moral cognition evolves by trial–and–error. It can be argued that inclusive-fitness evolution has developed the irreversible feature of moral beliefs to govern competition between individuals and groups, in adaptation to the ‘trial–and–error’ feature of this cognitive field.
The question now must be asked of how beliefs and the corresponding feelings/emotions are formed. At the behavioral level, beliefs are ‘imprinted’2 in a universal behavioral program, which can be called ’the human behavior instinct’ (it is similar to the universal grammar program, or ‘the language instinct” (PINKER 1994), where words are imprinted). The phylogenetically evolved program is receptive to, and will recognize, or ’learn’ the cultural instructions for behavior provided by group members. At the neurophysiological level, the beliefs correspond to the formation of ‘somatic markers’ postulated by Antonio DAMASIO. 3 Peer groups (HARRIS 1998) provide the ‘learning’ of social behavior, and this is one more example of how the behavior of groups, and of the individuals that compose them, is one–and–the–same phenomenon. It can be predicted that further neuropsychological investigations will reveal the existence of two kinds of somatic markers: reversible ones, corresponding to objective cognition (heteronomy) and to KANT’s ‘hypothetical imperatives’, and irreversible ones, corresponding to moral cognition (autonomy) and to KANT’s ‘categorical imperative’.
In conclusion, I would like to state again that in my opinion a more thorough explanation of human behavior can only arise from a comprehensive interdisciplinary approach, in which all the strata of the
Author’s address
Carlos Stegmann, Rua dos Pessegueiros, 101; 05673 – 010 Sao Paulo, Brazil. Email: stegmann@amcham.com.br
integrated natural entity Homo sapiens are considered. It is my considerate belief that only such an approach can provide a compass to navigate in the dark sea of human brain and mind phenomena.
Notes
with organisms which possess this faculty. The mechanism may likewise be extended to human beings, who may be said to be imprinted by the conditions of their civilization.” (The “conditions of their civilization” are imprinted as beliefs in the human mind.)
3 Antonio DAMASIO, with his theory of “somatic markers” (DAMASIO 1994; DAMASIO et al. 1994), has proposed the mechanism of the interaction between objective cognition and decision-governing feelings/emotions (beliefs). The referred works are not included in PANKSEPP’s references. The interdisciplinary interpretation (reduction) of DAMASIO’s findings (they were confirmed by neuropsychological tests) follows from the interdisciplinary relations extended order → individual behavior → somatic markers.
Evolution and Cognition ❘ 23 ❘ 2001, Vol. 7, No. 1
References